您好,欢迎访问三七文档
当前位置:首页 > 商业/管理/HR > 经营企划 > 异常重组转座遗传因子
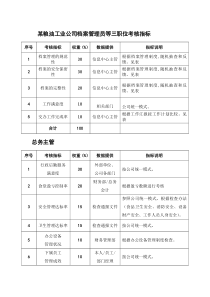
异常重组——转座遗传因子遗传重组异常重组的类型原核生物中的转座因子真核生物中的转座子转座因子的遗传学效应及其应用遗传重组同源性重组如真核生物在减数分裂时期同源染色体的非姐妹染色单体之间所发生的重组位点专一性重组如l噬菌体DNA通过其att位点和大肠杆菌DNA的attB位点之间专一性重组而实现整合过程。异常重组如转座子从染色体的一个区段转移到另一个区段或从一条染色体转移到另一染色体前两者可称为同源重组,异常重组也称为非同源重组。位点专一性重组(site-specificrecombination)1、概念:发生在专一序列的DNA分子间的重组,有特异的重组酶和辅助因子对其识别和作用。噬菌体基因组整合到细菌染色体基因组中属此种重组2、位点特异性重组的结果依赖于重组位点的位置和方向二、λphage的整合与切除1、实现机制:均是通过---细菌DNA和λDNA上特定位点之间的重组2、特定位点-----附着位点(attachmentsiteatt)◘E.coliattB含BOB’序列23bp◘λphageattP含POP’序列240bp◘核心序列“O区”完全一致(同源部分15bp)---位点特异性重组发生的地方3、整合过程◘整合后的附着位点为attL(BOP’)attR(POB’)◘整合位点---attB、attP切除位点---attL、attR◘整合过程需要λ整合酶(integraseInt)(λ编码)和寄主的整合宿主因子IHF(integrationhostfactor)共同作用溶源性细菌(lysogen)溶菌周期(lysis)原噬菌体概念及类型转座重组:转座子通过转座酶改变自己位置所引起的重组。转座子(transposon,Tn):能通过转座酶改变自己位置的DNA序列。转座子转座时插入的位点被称为靶位点。异常重组:是指不依赖于同源序列就能改变特定DNA序列位置,引起遗传性状改变的重组。以转座重组较普遍。根据转座中转座子复制与否,可分为两种:(1)复制型转座:在转座过程中转座子被复制,1个拷贝保留在原位点,1个拷贝被转移到新位点。依赖于转座酶及解离酶。(2)保守型转座:转座过程中转座子作为一个实体被转移到一个新位点。最早是McClintock(1951)在玉米粒色遗传研究中发现的Ac-Ds转座系统。基因水平转移水平基因转移(horizontalgenetransfer,HGT),又称侧向基因转移(lateralgenetransfer,LGT),是指在差异生物个体之间,或单个细胞内部细胞器之间所进行的遗传物质的交流。差异生物个体可以是同种但含有不同的遗传信息的生物个体,也可以是远缘的,甚至没有亲缘关系的生物个体。单个细胞内部细胞器主要指的是叶绿体、线粒体及细胞核。水平基因转移是相对于垂直基因转移(亲代传递给子代)而提出的,它打破了亲缘关系的界限,使基因流动的可能变得更为复杂。转座因子在原核生物中首次发现和检出BMcClintock通过对玉米花斑糊粉层和植株色素形成的遗传研究,首次清楚地表明,在任何基因组中都存在可移动的DNA序列(movableDNAsequences),但首次分离和检出这些可移动序列是在E.coli中。一、原核生物转座子二、真核生物转座子(一)插入序列IS;(二)转座子;(三)转座噬菌体(一)玉米转座子:Ac-Ds系统(二)酵母转座子:Ty因子(三)果蝇转座子:P因子(四)人类的转座子:LINE-1(L1)一、原核生物中的转座子类型按分子结构及遗传性质分类:插入序列(insertedsequence,IS):序列较小,含有转座酶等相关基因,两端有几个到几十个bp构成的反向重复序列。转座子(transposableelements,Tn):分子较大,除了含转座相关基因外还含有抗药性基因等其它基因,其两端有反向重复序列。转座噬菌体(transposablephage):可随机整合进细菌DNA任何位点的特殊噬菌体。分子大。(一)、插入序列(IS,insertedsequence)IS1•插入序列是最简单的转座因子,IS是细菌染色体和质粒的正常组成部分,可进行同源序列间的重组。E.coliK12chromosome8个IS1,5个IS2和IS3。IS两端的反向重复序列(二)转座子(transposon)在IS序列基础上,多了一些额外的基因如抗性基因。目前已发现40多种,如Tn1、Tn2、Tn3、Tn10等。Tn3的结构模式图:复合转座子有时两个IS序列之间虽然间隔有一些其它基因如细菌染色体基因、抗性基因等,但是它们可作为一个整体进行转座,改变位置。这种转座子被称为复合转座子(即带有已知的IS的Tn)。最早发现的是:1963年Taylor发现的是Mu-phage(Mutatorphage)。特点:1、是一种以大肠杆菌为寄主的温和噬菌体,以裂解生长和溶源生长两种方式交替繁衍自已,同时它又能像IS和Tn一样可以在宿主基因组上随机进行转座。即Mu噬菌体具有温和噬菌体和转座因子的双重特性。2、结构特点:Mu是一种DNA噬菌体,38000bp线状DNA;有三个区:α区:含有包括A、B基因等大多数基因;右侧3kb的G区序列:含Sv、U、U′和Sv′4个基因β区:含gin等基因。3、游离:两端连接着一段寄主DNA,左端100bp,右端1500bp;整合:再一次整合时,这两段序列消失(三)转座噬菌体在转录时G区序列的不同走向导致了不同的寄主特异性:G(+)→Sv和U基因表达→吸附E.coilK12菌株G(-)→Sv′和U′基因表达→E.coilC菌株转座噬菌体结构模式图二、真核生物中的转座子酵母菌是低等真核生物,其转座子类似于细菌转座子。酵母转座子中研究较清楚的是TY(transposonyeast)类转座子:如TY1和TY917。TY结构:含约5.6kb中心区,分布于两端的340bp的同向重复序列(称为δ),其作用与IS、Tn中的反向重复序列类似)。每个单倍体酵母基因组有30-35个TY,及至少100个单一的δ因子(soloδelements)。(一)酵母菌的转座子酵母Ty1转座子的结构(a)与Soloδ的形成(b)Ty1因子转座是通过一种RNA中间产物进行的。首先以其DNA为模板合成一个拷贝的RNA;然后再通过反转录合成一条新的Ty1因子;最后这条新的Ty1转座子再插入到新的位点上。Ty1因子转座(二)果蝇转座子果蝇中也有很多转座子:如Copia、P、412、279、Tip、FB等。其中,P因子可非复制型转座插入W位点,引起杂种劣育。例如卵巢发育异常,分离比异常,雄性个体的减数分裂中出现重组,突变率高,染色体畸变等。P因子:2907bp,两端为31bp的反向重复序列,中间含4个编码区(ORF0,ORF1,ORF2,ORF3)和3个内含子(1,2,3)。在体细胞中,内含子1、2被剪接掉,所形成的mRNA翻译成一个转座阻遏蛋白,抑制P因子转座。在生殖细胞中,内含子1、2、3都被剪接掉,所形成的mRNA翻译成转座酶,导致P因子转座,插入W位点引起配子劣育。在转座子切离时可以是准确的,也可能不准确,准确的切离,导致插入位点所在基因的回复突变,即恢复功能。不准确的切离,导致插入位点基因的突变,由此发生的突变有白眼、焦刚毛、黄体等。黑腹果蝇中的P因子与杂种败育在黑腹果蝇有两类品系:P品系:作为父本能造成杂种败育。其P因子含有正常转座酶基因(相当于显性不育基因S),但细胞质正常,有转座阻遏蛋白,能抑制转座(相当于细胞质中有该显性不育基因的抑制基因I)。基因型可表示为I(SS)。正常可育。M品系:作为母本能造成杂种败育的品系。其P因子缺失了编码区(相当于核内没有S),其细胞质中没有转座阻遏蛋白(相当于细胞质中没有I)。基因型可表示为i(ss)。P×P,P×M:杂种正常,基因型:I(_)M×M:杂种正常,基因型:i(ss)×i(ss)=i(ss)。M×P:杂种败育。i(ss)×I(SS)=i(Ss)。因为M品系细胞质中没转座阻遏蛋白,P品系的P因子能表达转座酶,可自由转座,因此杂种败育。(三)玉米转座子美国玉米遗传学家McClintock(1940-1950)发现玉米子粒色斑的不稳定遗传现象。经长达10年的研究,提出Ac-Ds转座系统。首次提出了遗传因子可以移动的观点,1983年获得诺贝尔奖。玉米糊粉层颜色至少与A、C、R、Pr、I等5个基因有关。其中A为花青素;C代表颜色,决定颜色(紫色或红色)的发生;R为红色,在存在A、C基因时决定红色的形成;Pr是紫色,在存在A、C、R下决定紫色的产生;I是颜色的抑制基因,抑制颜色产生,位于C基因附近。P252I=DsI可以发生位置改变,它解离转座到别处时,由于没有它的抑制作用,A、C、R共同作用可产生红色,因此被称为Ds(dissociator)。当它停留在原位点时,子粒表现无色。在A-C-R或A-C-R-Pr基因型的胚乳发育过程中,I解离越早,子粒上色斑的面积就越大;解离越晚,则子粒上色斑面积越小。由于不同胚乳细胞中Ds解离的时间不同,因此在子粒上产生大小不同的色斑嵌合现象。Ac-Ds系统Ac也是一个转座子,由4563bp组成(其中间区编码转座酶),两端有11bp的反向重复序列,可转座到基因组的任何位置,其靶位点有两个8bp的同向重复序列。其中间编码区不同程度缺失,形成不同的Ds。当Ac开始转座活动时,Ds被激活也进行转座,移动到新位点,使靶位点附近基因失活或改变表达活性。•研究发现,Ds是激活因子Ac(activator)的缺失突变体,不能自主转座,需被Ac因子激活。尤其是两端重复序列与Ac同源。Ac-Ds的类似系统在玉米中,除了Ac-Ds系统外,还有其它5个转座系统。但是它们之间是类似的。三、转座机制和遗传学效应转座步骤是:(1)转座酶在靶位点上制造一个交错的切口。(2)然后转座子与突出的单链末端相连接,并填充缺口。交错末端的产生和修复,可产生靶位点的同向重复。其交错长度决定了同向重复的长度。如下图:(一)转座机制转座机制逆转录转座子DNA—RNA—DNA的转移过程——逆转录转座。逆转录子——一类移动因子在转座过程中需要以RNA为中间体,经过逆转录过程再分散到基因组中。在所有高等生物的基因组构成中,存在着与逆转录病毒基因组非常相似的逆转录转座子。逆转录转座子与简单转座子比较1.与高频率自发突变有关;2.刺激寄主细胞基因组发生多种形式的遗传重排;3.插入单元两端的DNA序列在插入过程中增加3-13bp;4.转座单元的末端含有长度为2-50bp的反向重复5.转座过程经常与转座单元的复制同时进行;6.转座子通过编码蛋白质作用于转座单元来对自身的转座功能进行调控。逆转录子的结构特点逆转录子编码逆转录作用的关键酶:逆转录酶和整合酶。按结构特征分两类(1)与逆转录病毒类似的LTR、gag和pol基因,但无env基因。(2)不具有LTR,但有3‘polyA、gag和pol基因。酵母果蝇哺乳果蝇(二)转座的遗传学效应1、引起插入突变(引起基因失活)或切离引起回复突变等。从而对基因表达进行调节。2、给靶位点带来新的基因,如转座子上的抗药性基因等3、引起序列重复:如转座子复制重复,靶位点同向重复序列。4、引起染色体结构变异:如处在不同位点(甚至不同染色体上)的同源转座子间可相互重组,导致染色体片段的重复、缺失、倒位等结构变异。如重组的两个转座子的重复序列相同,重组后产生缺失;如果相反,则重组后发生倒位。5、可能诱发新的变异,产生新基因。转座时可引起插入突变。6、转座子在作为基因转移的供体或标记,在遗传研究及基因工程等方面也有广泛的用处。
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码









 jeromeziv
jeromeziv
本文标题:异常重组转座遗传因子
链接地址:https://www.777doc.com/doc-2429002 .html