您好,欢迎访问三七文档
当前位置:首页 > 建筑/环境 > 工程监理 > 基因工程 宿主与转化
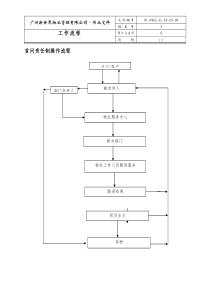
生物技术第四章基因表达系统基因表达系统4.1植物基因转化4.2原核表达系统4.1农杆菌介导的植物基因转化一、农杆菌农杆菌:根瘤菌科(Rhizobiacease);土壤杆菌属(Agrobacterium),革兰氏阴性。根癌农杆菌(Agrobacteriumtumefaciens)放射型农杆菌(Agrobacteriumradiobacter)发根农杆菌(Agrobacteriumrhizogenes)悬钩子农杆菌(Agrobacteriumrubi)农杆菌是植物的病原菌根癌农杆菌感染导致冠瘿瘤的形成Themechanismofcrowngallformationhasbeenextensivelyreviewedbyanumberofauthors(Zupanetal.,2000;ZupanandZambryski,1995;Zambryski,1992;ShengandCitovsky,1996;HooykaasandBeijersbergen,1994).植物病原菌,天然遗传转化系统载体系统-Ti质粒宿主范围广泛存在于双子叶植物中,据不完全统计,约有93属643种双子叶植物对根癌农杆菌敏感。裸子植物对该菌也具有敏感性,能诱发肿瘤。一般认为单子叶植物对农杆菌无敏感性。DeCleeneandDeLay(1976)reportedalistofnaturalhostplantsforAgrobacteriumandtheirlistcontained42gymnosperms,596dicotyledonsand5monocotyledons.Thenumberofmonocotyledonsinthelistwasverysmallandnoagronomicallyimportantcerealswereincluded.Thus,thegeneralstatementthatAgrobacteriumdoesnotinducetumoursonmonocotyledonsisnotliterallytrue,butitreflectsgeneralbeliefsoveralongperiodoftime.单子叶植物与双子叶植物对损伤的响应不同,对宿主范围有影响3种类型-用损伤细胞作为保护层(保护层内的细胞木质化)单子叶植物-细胞分裂,形成愈伤组织双子叶植物-形成创伤周皮二、冠瘿瘤形成机制Ti质粒编码蛋白介导T-DNA转移;T链(单链)进入植物细胞核,整合到植物基因组中1.Ti质粒双链共价闭环DNA,分子量约200kb迄今已从多种植物分离了不同种类的农杆菌,它们的Ti质粒的,结构特性均已清楚。菌株起源致瘤质粒名称B6苹果瘤+pTi-B6209番茄瘤+pTi15955NCPPB1001葡萄瘤+pTi1001C58樱桃瘤+pTi-C58pAT-C5827917长病桃树的土壤+pTi-1D13527白杨瘤+pTi-27EU6卫矛瘤+pTi-EU6pAT-EU6Ti质粒的功能区T-DNA整合到植物染色体Vir基因介导基因转化Ti质粒复制起始区Vir区T-DNA区冠瘿碱代谢区1982,Chilton等人用同位素标记Ti质粒做探针与烟草冠瘿瘤细胞DNA杂交;后将章鱼碱型Ti质粒用内切酶SmaI切成19个片段,分别制备探针后与冠瘿瘤DNA杂交;研究结果:两段Ti质粒DNA能和冠瘿瘤DNA杂交,说明这部分DNA从Ti质粒上切割后转移到肿瘤细胞中LBRBAuxiniaaMiaaHtmsCytokininipttmrOpinetmt冠瘿碱代谢区复制起始区Vir区Ti质粒tms的编码产物负责合成吲哚乙酸tmr的编码产物负责合成细胞分裂素tmt的编码产物负责:合成氨基酸衍生物T-DNA的结构左右边缘序列编码基因可以在植物细胞中表达T-DNATms和Tmr基因的表达使植物细胞内激素平衡紊乱,冠瘿瘤细胞无限生长,形成激素自主的特性,导致冠瘿瘤形成。Tms和Tmr基因是致瘤所必需的基因,称为致瘤基因(oncgene)Vir区基因的功能-V区基因介导T链转移-至少6个操纵子在链转移中起作用virA(1cistron),virB(11cistrons),virC(2cistrons),virD(4cistrons),virE(2cistrons)andvirG(1cistron)(StachelandNester,1986).-这些基因在侵染植物时表达2.转化机制信号受体VirA识别化学信号,通过磷酸化激活VirG;VirG为转录调节因子,可上调V区基因表达损伤后植物体的汁液呈酸性,含各种物质,包括木质素,类黄酮生物合成途径中产生的酚类物质,受损伤后的植物汁液诱导v区基因表达(Stacheletal.,1985),其中最有效的酚类化合物是乙酰丁香酮T链的形成及转运在VirD1,VirC1(and,possibly,VirD3)蛋白的协助下,VirD2蛋白识别左、右边缘序列,在第3和第4个核苷酸之间切割下面单链产生缺口(Zambryski,1992).VirD2共价结合T链5’端,随着Ti质粒DNA的合成,T链被“释放”。VirE2为单链DNA结合蛋白,VirE2结合T-DNA形成T链复合体;T链的转化仅要求边缘序列VirD2VirB和VirD4是膜蛋白,在农杆菌与植物细胞之间形成通道,T链复合体经该通道进入植物细胞质,从植物细胞质到植物细胞核由VirD2好VirE2蛋白的核定位信号介导(ShengandCitovsky,1996).整合T链复合体进入植物细胞核后,会形成双链,T-DNA编码的基因在整合前会表达,被称为瞬时表达(Kapilaetal.,1997).不清楚双链的形成发生于整合前还是整合后,人们认为整合从左边缘开始(Konczetal.,1994).整合可能由宿主DNA复制和修复机制中的酶介导,插入位点是随机的(Konczetal.,1994),有人提出VirD2andVirE2可能参与整合(Rossietal.,1996).附着细菌附着植物细胞壁是转化的前提条件;农杆菌从植物伤口进入植物体,在植物体汁液中繁殖并附着到植物细胞壁上(Zambryski,1992),有学者认为农杆菌染色体编码的基因chvA、chv、pacA、att和植物细胞表面受体(包括蛋白和糖)参与细菌的附着(ShengandCitovsky,1996)ChvA、ChvB、ChvC农秆菌附着功能有关,这三个基因突变使农杆菌与植物细胞的附着率下降10倍;ChvB基因产物催化合成β-1,2-葡聚糖,由ChvB调控的β-1,2-葡聚糖可能在农杆菌附着过程中起关键作用;ChvA编码产物与多糖从细胞质转运胞外有关。ExoC与环化葡聚糖和琥珀酸多聚糖的合成有关,ExoC基因突变,农杆菌不能合成环化的β-1,2-葡聚糖,细菌不能附着在植物细胞上,无致瘤能力。Cel与细菌表面纤维丝的合成有关,Cel基因突变体不能合成纤维素细丝,细菌附着到植物细胞表面后容易被水洗脱。Att影响农杆菌细胞表面蛋白的合成,Att突变体缺少一种34kD和38kD的多肽,这些多肽是细胞外膜和细胞周质蛋白,影响农杆菌附着。三、植物基因表达载体tmrtmstmtT-DNALBRBvirTi质粒野生型Ti质粒的缺点:•Ti质粒分子量过大,难操作;•Onc基因致瘤onc改造:•除去ONC基因•安装大肠杆菌复制子;•安装植物细胞选择标记一元载体系统也称共整合载体(co-integratedvector),顺势载体(cis-vector)卸甲载体中间载体一元载体系统卸甲载体:切除T-DNA中的Onc基因,称disarmedvector;作用:提供vir功能virori冠瘿碱分解代谢LB卸甲Ti质粒经典卸甲载体:pGV3850T-DNA边界区,胭脂碱合成酶基因(nos);T-DNA以外的全部序列;T-DNA缺失部分被pBR322序列(含Ampr)取代pBR322nosvirpGV3850中间载体(intermediatevector)大肠杆菌的克隆载体(如pBR322),应是多拷贝小质粒。特征:•含与卸载质粒T-DNA区同源的序列,可与卸载质粒T-DNA高频地同源重组;•具有细菌选择标记,便于筛选共整合质粒;•有bom位点,便于结合转移;•含有植物选择标记,如新霉素磷酸转移酶基因(Npt-II);•含单一的限制性内酶位点;•植物的启动子。作用:用于克隆目的基因;通过同源重组机制将目的基因整合到卸甲载体的T-DNA上;提供T-DNA边缘序列共整合转化程序基础:农杆菌与大肠杆菌之间可以高效地接合转移;Tra基因群功能T-DNApBR322oriNPTII(Kmr)大肠杆菌质粒大肠杆菌筛选标记农杆菌筛选标记多克隆位点重组质粒转化大肠杆菌农杆菌细菌接合T-DNA区同源整合农杆菌感染植物根部细胞T-DNA区整合在植物细胞基因组上三亲株杂交(triparentalmating)通过细菌结合转移机制将中间载体导入农杆菌农杆菌菌株:受体;卸甲Ti质粒;农杆菌复制起点大肠杆菌:辅助质粒;提供Tra功能和Mob功能;大肠杆菌复制起点大肠杆菌:中间载体;穿梭质粒;bom位点;E.coli复制起点、广谱复制起点pRK2013Tra,mob+E.coliKmE.colipMon200bom+Spc/StrpRK2013pMon200pTiB6S3-SEbom+Spc/StrKmLBRBYFG农杆菌农杆菌三亲株杂交三亲株杂交TriparentalMatingSpc壮观霉素Str链霉素Km卡那霉素pRK2013—结合转移型质粒,E.coli复制子pMON200—中间载体,E.coli和农杆菌复制子双元载体系统T-DNA与Vir基因位于两个独立的质粒上,通过反式作用激活T-DNA转移,称反式载体(transvector)辅助质粒virLBRB微型Ti质粒微型Ti质粒(mini-Tiplasmid):T-DNA边界序列广谱质粒复制位点,如RriV原核和真核选择标记克隆位点辅助质粒(helperplasmid):缺失T-DNA区段的Ti质粒,本身丧失致瘤功能,相当于卸载Ti质粒。主要作用是提供Vir基因功能。经典双元载体Bin19:pTiT37的左右边界序列;Npt-II基因;LacZ基因;多克隆位点;广泛宿主质粒Rk2的复制和转移的起始位点优点:质粒小、宿主广;带有EcoRI、BamHI、HindIII、SstI、KpnI、SmaI、XbaI及SalI组成的多克隆位点;可以进行蓝白斑筛选;可以直接转化农杆菌(冻融法)二元整合转化程序二元整合转化程序LBRBT-DNA外源基因植物细胞筛选标记Kmr大肠杆菌-农杆菌穿梭质粒大肠杆菌筛选标记农杆菌筛选标记农杆菌ori大肠杆菌ori将外源基因克隆在大肠杆菌-农杆菌穿梭质粒的T-DNA区内;将外源基因克隆在大肠杆菌-农杆菌穿梭质粒的T-DNA区内;重组质粒直接转化农杆菌株,该菌株携带只含vir区不含T-DNA区的Ti辅助质粒;重组质粒直接转化农杆菌株,该菌株携带只含vir区不含T-DNA区的Ti辅助质粒;以上述重组农杆菌感染植物细胞。以上述重组农杆菌感染植物细胞。四、植物基因转化程序基因构建农杆菌菌株受体植株组培体系植物基因转化植物基因转化程序培养纯化农杆菌农杆菌菌液预培养植物材料植物材料共培养→抑菌和选择培养↓芽分化↓再生植株分子鉴定表型筛选田间试验遗传稳定性研究感染1.基因构建启动子目的基因Nos3`LBMCSnosPNpt-IInos3’RBBin19微型Ti质粒EcoRIKpnISmaIBamHIXhaISatIHindIIILB启动子目的基因Nos3`nosPNpt-IInos3’RB大肠杆菌克隆重组质粒农杆菌工程菌株重组微型Ti质粒转化测序鉴定转化农杆菌卸甲Ti质粒LBPNptIIterP目的基因terRBvir农杆菌重组微型Ti质粒-70℃20%甘油保存植物基因转化前期研究中,学者们构建了一些载体,主要是双元载体(HellensandMullineaux,2000)。这些载体的应用不依赖于
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码







 wz000140
wz000140
本文标题:基因工程 宿主与转化
链接地址:https://www.777doc.com/doc-4069489 .html