您好,欢迎访问三七文档
当前位置:首页 > 医学/心理学 > 药学 > 肿瘤干细胞的研究进展
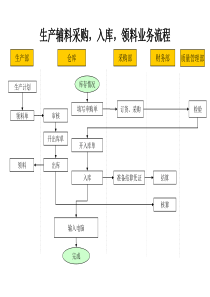
1肿瘤干细胞的研究进展Researchadvancesoncancerstemcell姓名:刘小阳学号:s0910260日期:2009/122Contents干细胞与肿瘤干细胞1肿瘤干细胞的起源2靶向肿瘤干细胞的研发热点3肿瘤干细胞理论存在的质疑43干细胞与肿瘤干细胞干细胞在细胞的分化过程中,细胞往往由于高度分化而完全失去了再分裂的能力,最终衰老死亡。机体在发展适应过程中为了弥补这一不足,保留了一部分未分化的原始细胞,称之为干细胞(stemcell)。干细胞的分类干细胞按分化潜能的大小,可分为三种类型:一类是全能性干细胞,它具有形成完整个体的分化潜能。另一类是多能性干细胞,这种干细胞具有分化出多种细胞组织的潜能,但却失去了发育成完整个体的能力,发育潜能受到一定的限制。4干细胞与肿瘤干细胞还有一类单能干细胞(也称专能、偏能干细胞),这类干细胞只能向一种类型或密切相关的两种类型的细胞分化。肿瘤干细胞把小鼠白血病细胞移植到同品系的小鼠体内,发现移植细胞仅有1%~4%能够形成脾脏内克隆,因此得出结论:只有部分肿瘤细胞(而不是全部肿瘤细胞)具有致瘤性。近年的研究显示,某些白血病仅有百万分之一的细胞具有这种无限增殖的潜能,它们数量虽少,但具有干细胞特征,可能是惟一保持增殖的肿瘤细胞,这就是肿瘤干细胞(cancerstemcell/CSC)。5干细胞与肿瘤干细胞这些干细胞也可以持续增殖分化,形成相对成熟的肿瘤细胞,它们不能再增殖,不能形成细胞集落,或者肿瘤细胞仅能分裂几次,体外克隆培养只能形成几个或几十个细胞,经短暂的分化最终死亡。而其中的肿瘤干细胞才能持续增殖,形成细胞集落,它们在肿瘤的形成和生长过程中起决定性作用,是肿瘤不断生长和扩大的源泉。2001年,Nature上发表文章提出当代肿瘤干细胞学说,认为肿瘤组织存在一小部分具有干细胞性质的肿瘤干细胞,其具有无限的自我更新、多向分化潜能与高度增殖能力,能够产生不同表型的肿瘤细胞,并赋予肿瘤异质、增殖、浸润等特性。随后,对于这个假说的验证试验进行得如火如荼。6图1肿瘤发生的CSC模型7干细胞与肿瘤干细胞干细胞与肿瘤干细胞的相似点两者均具有自我更新和无限增殖能力。干细胞与肿瘤干细胞均有较高的端粒酶活性。有不同分化程度的干细胞,也有分化程度不同的肿瘤。干细胞的迁移和肿瘤细胞的转移都有一定的组织和器官特异性,且二者皆受特异性化学因子及其受体的调节。二者有许多相似的生长调控机制,如Wnt,Sonichedgehog,Notch途径。自我更新越频繁的组织,干细胞自我更新率越快,肿瘤的发生率越高。具有比较类似的表面标志物。8干细胞与肿瘤干细胞干细胞与肿瘤干细胞(CSC)的区别干细胞可以在某一时间内连续分裂,也可在较长时间内处于相对静止状态,干细胞的自我更新具有反馈机制调节,增殖和分化处于平衡状态,是有序的;而在肿瘤干细胞中,这一调节机制并不存在,它的增殖分化是无序的,失控的。肿瘤干细胞没有分化为成熟细胞的能力,其分化程序异常,这与有正常分化程序的干细胞有着本质的不同。肿瘤细胞倾向于积累复制错误,而正常干细胞的发育机制要防止这种现象的出现。9肿瘤干细胞的起源许多研究支持CSC来源于正常干细胞目前大多数学者支持来源于正常干细胞的观点,认为肿瘤的发生是因为体内干细胞分化过程的停滞和阻断,细胞失去正常调控而停止在分化的某一阶段无限增殖形成,而不是成熟细胞的去分化。由于干细胞自我更新能力已经激活,少量突变持续发生就会导致细胞癌变,使干细胞容易成为恶变的靶细胞。白血病初始细胞表型为CD34+CD38-,研究表明Thy-1(CD90)的丢失导致了这种恶性转化。10肿瘤干细胞的起源已经证实白血病干细胞不表达c-Kit(CD117),但表达IL-3Rα(CD123),因此白血病干细胞可能是由造血干细胞衍生而来。也有研究表明CSC不仅可起源于正常干细胞,也可起源于祖细胞,如骨髓祖细胞。由于干细胞在个体生长发育过程中不断增殖和自我更新,因此发生突变的几率更大。当受到各种致癌因子作用或所处微环境改变时,不断增殖的细胞发生突变并累积,突变发生在正常干细胞和祖细胞,均能形成肿瘤(图2所示)。11图2肿瘤干细胞起源示意图12肿瘤干细胞的起源肿瘤干细胞起源于组织内干细胞异常分化的理论依据一个正常细胞发生恶性转化至少要4~7次突变。由于自然情况下突变是一个低概率事件,要积累如此多的突变需要几年甚至几十年时间。在此期间,只有干细胞能自我更新而一直存在。干细胞无限增殖的特性增加了其发生增殖错误的机会。干细胞由于连续复制,能够将变异的基因遗传给子代细胞,造成变异的基因在子代细胞中积累而导致细胞恶性转化。干细胞及其前体细胞已具备活化的自我更新机制,显然维持这一机制要比在已分化细胞中重新激活这一机制要容易。肿瘤干细胞与组织干细胞有相似的生长调控机制。肿瘤干细胞具有许多干细胞共同的特异性标志物。13靶向肿瘤干细胞的研发热点靶向肿瘤干细胞特有标记利用携带细胞毒药物的抗体靶向相应的肿瘤干细胞特有标记,或找到与其他种类细胞不同的参数来区分出肿瘤干细胞,进而特异地消灭这一细胞亚群。但由于肿瘤干细胞在肿瘤组织中存在比例很小(小于5%)以及目前经实验鉴定出的肿瘤干细胞表面标记还十分有限,并且通过分析细胞表面标记来鉴定其中干细胞的方法对实际应用来说太过复杂,所以利用携带药物的抗体靶向特殊表面标记存在很大的困难。目前,根据特异性标记从多种肿瘤中分离出肿瘤干细胞,并对其进行了分析和鉴定(表1所示)。14表1不同种类癌症中肿瘤干细胞的标志物15靶向肿瘤干细胞的研发热点当前针对靶向肿瘤干细胞特异性标记的研究,主要是表面标志物的寻找及内部特异性蛋白的鉴定。目前发现了一种能用于鉴定乳腺肿瘤中干细胞的标志物,乳腺癌中ALDH(醛脱氢酶)阳性的肿瘤细胞具有自我更新的干细胞特性,并且500个这样的细胞便能形成一个肿瘤,相反,ALDH阴性的细胞不具有这样的特性。此外还在乳腺肿瘤中干细胞确认了一种具有11个基因的hSAGA复合物可以作为癌干细胞的标记物,而且USP22作为此复合物中的一部分对细胞的转录具有重要的作用,将成为癌症治疗的靶标。16靶向肿瘤干细胞的研发热点最近报道了美国俄克拉荷马大学癌症研究院的一项研究成果,该机构发现一种仅在癌症干细胞中存在的蛋白,并对靶向该蛋白的化合物进行筛选。研究人员表示如果该药物能够通过人体实验,有望在十年内开发出针对肿瘤干细胞的新药。靶向肿瘤干细胞微环境肿瘤干细胞与周围微环境之间有着千丝万缕的联系,例如受众多细胞因子、配体的调控,以及各种类型细胞的支持等,以保持其在肿瘤组织中自我更新的特性(图3所示)。通过靶向调控肿瘤干细胞的微环境,使其丧失自我更新的功能,进而切断肿瘤发展的源泉。目前微环境对肿瘤干细胞的调控机制知之甚少,但最近的一些研究结果对于靶向肿瘤干细胞微环境提供了有力的依据。17图3肿瘤干细胞的微环境18靶向肿瘤干细胞的研发热点肿瘤干细胞作为肿瘤转移的元凶,受到周围炎症环境的影响,因此靶向炎症因素可以防止肿瘤的转移。针对胰腺癌的研究表明,趋化因子受体CXCR4能够赋予肿瘤干细胞转移的特性,在去除CXCR4后,肿瘤干细胞只具有致瘤性而失去转移特性,这预示着CXCR4作为炎症环境中的靶点对防治肿瘤转移具有重要的意义。最近的一项研究进一步阐明了肿瘤转移的微环境机制,结果表明肿瘤细胞受到S100A8和S100A9等趋化因子的诱导向肺部转移,同时诱导肺部细胞分泌一种血清淀粉样蛋白A3(serumamyloidA3,SAA3),SAA3通过Toll样受体(Toll-likereceptor,TLR)激活NF-kB通路,加速肿瘤细胞的募集并进一步促进趋化因子的分泌(图3所示)。因此,靶向抑制TLR4或SAA3能够降低小鼠肺部肿瘤的转移。19靶向肿瘤干细胞的研发热点靶向肿瘤干细胞关键通路肿瘤干细胞与正常干细胞在自我更新方面有着相似的信号通路包括Wnt/b-catenin、Notch、Hedgehog等(图4所示),这些通路在正常干细胞中的紊乱可以导致肿瘤的发生。2008年10月,肿瘤测序计划(tumoursequencingproject,TSP)的研究结果,其显示Wnt作为维持干细胞自我更新的重要通路在肺癌中的突变率达到15℅,其中担任重要角色的蛋白APC、CTNNB1、GSK3b等均发生了显著的突变。因此,靶向肿瘤干细胞中异常的维持自我更新的通路对消灭肿瘤有着重要的意义。20图4肿瘤干细胞主要通路及靶向图示21图4中三条信号通路说明如下:•Hedgehog信号通路:Hh配体与Pct受体结合,解除了Pct受体对膜蛋白Smo的抑制作用,后者促使锚定在微管上的Cos复合物脱离微管,将有正调节活性的锌指转录因子(Ci)从中释放出来,进入细胞核诱导目的基因的表达。•Wnt/β-catenin信号通路:当Wnt配体作用于受体及辅助受体后,激活Dvl蛋白,进而解除了“蛋白降解复合物”(GSK3b-APC-Axin)对β-连环蛋白(β-catenin)的降解,使其进入细胞核与转录因子LEF/TCF等作用激活靶基因转录。•Notch信号通路:当Notch受体被相邻细胞的Delta/Jagged配体激活后,分别经过由肿瘤坏死因子-转化酶(TNF-aconvertingenzyme,TACE)以及γ-分泌酶(γ-Secretase)两次剪切,在胞内释放Notch蛋白的活化形式—NICD,NICD进入核内与转录因子作用诱导靶基因的表达。22靶向肿瘤干细胞的研发热点靶向肿瘤干细胞关键通路通过靶向阻断脑瘤细胞中Notch通路,发现肿瘤细胞中凋亡及分化数量明显增高,而分化的肿瘤细胞不再具有致瘤特性。阻断Notch通路能够明显减少CD133+脑肿瘤干细胞的数量并促进nestin+细胞的凋亡。最近的研究表明,应用γ-分泌酶(γ-Secretase)抑制剂(GSIs)以阻断Notch通路(图4所示),可以抑制乳腺癌干细胞的生长并消除其自我更新功能,并有望在今后的临床实验中获得成功。23靶向肿瘤干细胞的研发热点分别用环杷明(Cyclopamine)和慢病毒载体LV-shSMOH作用于胶质瘤细胞,用以阻断维持CD133+胶质瘤干细胞自我更新的Hedgehog-GL1通路(图4所示)。结果表明经环杷明处理20d的胶质瘤球全部被杀死,且经转染慢病毒载体的细胞也受到明显抑制。而用DNA损伤剂TMZ(替莫唑胺)处理的细胞,由于不能杀死其中的肿瘤干细胞故效果不如前两者。24CSC理论存在的质疑克隆进化理论的挑战人们对于肿瘤发生与发展的认识除了肿瘤干细胞模型以外,主要还存在一种与之相对的理论,即克隆进化模型。该模型认为肿瘤细胞是经过长期的克隆演变进化而来,肿瘤起源于正常的细胞,这些细胞突变后产生了异常的子代细胞克隆,而这些子代细胞继续突变,如此经过数代的突变累积,最终产生大量遗传变异的癌细胞并促成了肿瘤组织的异质性。25CSC理论存在的质疑来自不同实验室的研究结果均对肿瘤干细胞模型提出了质疑,而偏向于支持克隆进化模型。通过基因扫描等技术,对人类乳腺癌样本中CD44+与CD24+细胞之间进行分子以及表型的分析,结果表明这两种细胞在基因表达水平及表观遗传方面并不相同,这与肿瘤干细胞理论所解释的CD24+细胞是CD44+细胞的后代的说法相悖,进而这种异质性与克隆进化模型更为相符。实验因素的质疑对于肿瘤干细胞理论的质疑,很大一部分是来自实验方面的因素。26CSC理论存在的质疑目前针对肿瘤干细胞检测的一个普遍的方法是异种异体移植实验,即具有免疫缺陷的非肥胖型糖尿病/重症联合免疫缺陷(NOD/SCID)鼠成瘤性测试,但这一实验方法近年来受到一些学者的质疑。研究者认为,异体移植实验掩盖了微环境对肿瘤细胞的影响,由于人与鼠体内的微环境的不同,不能够说明在鼠体内有致瘤性的那些来自人的癌细胞即为肿瘤干细胞。人的肿瘤细胞之所以能够在鼠体内存活并成瘤,可能是由于这些细胞得到了某些特性从而能够适应新的环境,而非这些细胞是具有自我更新功能的癌干细胞
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码








 互动户的嘎雅
互动户的嘎雅
本文标题:肿瘤干细胞的研究进展
链接地址:https://www.777doc.com/doc-4261986 .html