您好,欢迎访问三七文档
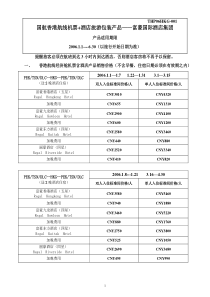
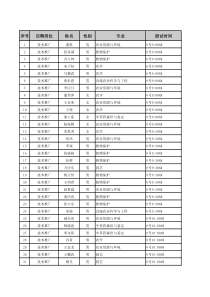
1第十一章肿瘤分子生物学2主要内容反转录病毒致癌基因1原癌基因产物及其分类2原癌基因的表达调控3基因相互作用与癌基因表达43环境致病因素如饮食、病毒、化学物质和射线的致癌作用都是通过遗传基因起作用癌基因即细胞中活化的原癌基因.对DNA复制、细胞生长与增殖起正相调节或促进作用抑癌基因即控制细胞生长、增殖及分化过程中起着十分重要的负相调节作用肿瘤是环境因素与遗传因素相互作用而导致的一类疾病45癌症的多阶段发生学说(multi-stage)6•癌(cancer):是一群不受生长调控而繁殖的细胞,也称恶性肿瘤。•良性肿瘤:是一群仅局限在自己的正常位置,且不侵染周围其它组织和器官的细胞。•绝大多数癌是由肿瘤细胞经过一系列突变转化而来的。7肿瘤细胞是永生化的、转化了的细胞细胞癌变时发生三种类型的改变:永生化(immortalization)细胞克服了细胞分裂次数的限制无限增殖的特性。转化(transformation)指在转化的细胞中,不能观察到正常的细胞生长受限制,如转化细胞不依赖于通常能够刺激细胞生长和生存的因子,细胞变圆并长成一个集落(实体瘤)。转移(metastasis)癌细胞获得了能够侵入正常组织的能力。它能够离开原来的组织,转移到机体的其他部分,并形成新的克隆。8异常生长的肿瘤细胞9WHO报道:•全球每年约有1000多万人新患癌症,•每年约有670多万人死于癌症,•几乎每5-6秒钟就有一名癌症患者死亡。我国目前:•每年平均约有150万人新患癌症,•每年约有80万人死于癌症,•癌症的死亡从1949年占总死亡原因的第十位已经上升到第一位。癌症:严峻的现实10•癌基因的发现以及研究的深入;•染色体畸变与致癌基因表达相互关系的揭示;•抑癌基因的发现及表达调控。肿瘤医学研究中最为突出的成就细胞癌变的分子基础是基因突变,DNA的变化和不正常活动导致了细胞癌变。11•癌基因(oncogene):是一类与肿瘤发生和发展密切相关的基因,在体外可促进细胞转化,在体内可诱导肿瘤的发生,其蛋白产物可促进细胞的增殖。•抑癌基因(tumorsuppressorgene,TSGs)抑制细胞生长。抑癌基因的活性下降(功能缺失突变)是引起癌变的另一个原因。12病毒癌基因(viraloncogene,V-onc):•致瘤病毒中能在体内诱发肿瘤并在体外引起细胞转化的基因•编码病毒癌基因的主要有DNA病毒和RNA病毒。•大约有15%-20%的癌症是由病毒感染引起的。细胞转化基因(cellularoncogene,C-onc):•存在于细胞基因组中,正常情况下处于静止或低水平(限制性)表达状态,对维持细胞正常功能具有重要作用,•当受到致癌因素作用被活化使正常细胞转化为肿瘤细胞。细胞转化基因与病毒癌基因有显著的序列相似性。癌基因(oncogene)可分为两大类:13•DNA病毒包括乙型肝炎病毒、SV40和多瘤病毒、乳头瘤病毒、腺病毒、疱疹病毒和痘病毒。•RNA病毒主要是反转录病毒。包括肉瘤病毒、HIV、流感病毒、冠状病毒等。反转录病毒致癌基因(retrovirusonc)可能是研究最多的病毒基因,它们能使靶细胞发生恶性转化。14与人类肿瘤发生有关的病毒移除肿瘤&制备无细胞滤液SarcomaSarcoma(一).反转录病毒致癌基因(Retrovirusonc)16(一)反转录病毒致癌基因Rous于1910年首先发现鸡肉瘤病毒(后称劳斯肉瘤病毒RSV),研究证明它是一种单链RNA肉瘤病毒(Roussarcomavirus,RSV,一种反转录病毒)1966年,因发现肿瘤诱导病毒,因提出病毒致癌学说,开创癌症起因研究的新纪元而获得1966年诺贝尔生理学医学奖。171.反转录病毒颗粒结构18调节和启动转录LTRgagpolenvLTR长末端重复序列正常的病毒基因产生病毒表面鞘蛋白产生逆转录酶和整合酶产生病毒外壳蛋白2.反转录病毒基因组结构19•基因组由单链RNA组成(6-9kb);•gag和env两个基因编码蛋白质外壳;•反转录病毒的复制是由其自身基因组上pol基因编码的反转录酶指导完成的2.反转录病毒基因组结构20当病毒感染细胞时,pol基因被宿主的RNA聚合酶II转录,产生polmRNA,并在宿主核糖体上合成包括反转录酶、整合酶等多个病毒复制和整合所需要的酶类。反转录酶以病毒RNA为模板,转录出单链DNA分子利用宿主DNA聚合酶指导合成第二条DNA链,以双链DNA形式整合到宿主细胞基因组中。21Baltimore1975年因为发现了逆转录酶,证明遗传信息不仅由DNA到RNA,也可由RNA到DNA而获得诺贝尔生理学医学获奖22•原病毒(provirus):被整合的反转录病毒DNA分子•它能指导病毒mRNA的合成,•并利用宿主细胞中的蛋白质合成机器,翻译生成病毒外壳蛋白等,最后组装成病毒颗粒。231970s发现RSV中的致瘤基因是src基因(最早研究的致癌基因),编码被称为p60Src的蛋白质。•是514个氨基酸的磷酸化蛋白(酪氨酸激酶),•其C端250个氨基酸是活性区域,负责将磷酸基团移到酪氨酸残基上,可使多个靶蛋白发生磷酸化而影响其功能,加速细胞癌变的过程。•p60Src结合在细胞基部质膜附着点内靠近或位于枢纽蛋白处,使该蛋白磷酸化水平提高20倍以上,导致细胞黏附能力减弱,易发生脱落和转移。24研究发现,反转录病毒基因组中所带有的onc基因并非来自病毒本身,而是这些病毒在感染动物或人体之后获得的细胞原癌基因。动物或人原癌基因经病毒修饰和改造后,成为病毒基因组的一部分并具有了致癌性,其作用的靶分子也往往发生改变。反转录病毒癌基因起源于细胞内的原癌基因3.反转录病毒癌基因(v-onc)的起源253.反转录病毒癌基因(v-onc)的起源26断裂基因完整的没有断裂的可读框肉瘤病毒V-Src基因,编码有514个氨基酸的p60Src磷酸化蛋白27284、根据反转录病毒转化细胞的能力,可分为:29除病毒感染诱发细胞癌变外,许多非病毒因子(如放射性物质、化学试剂亚硝酸、烷化剂等)也能诱导细胞转化。这些因子没有把致癌基因或其他致癌的遗传信息带入细胞,而通过某种激活机制改变了细胞内原有的遗传信息,使细胞发生恶性转化。→致癌因子导致基因突变!(二)原癌基因产物及其分类301、概念.原癌基因(pro-oncogene):正常细胞中存在癌基因,在正常情况下它们不具有致癌作用,但在一定情况下被激活后可变成具有致癌作用的癌基因,在正常情况下它们不但无害,而且对细胞的发育、生长和分化的调节起重要作用。31原癌基因只是为了使机体发生癌症而存在吗?322.原癌基因的特点:广泛存在于自然界在进化上高度保守是调节细胞生长、发育和分化的重要蛋白质在结构上与病毒癌基因相近正常情况下表达水平很低在一定条件下可被激活成癌基因33美国人M.Bishop与美国科学家Varmus因在70年代最终发现了细胞癌基因(原癌基因),创立了癌症发生的癌基因理论。获1989年诺贝尔生理医学奖。BishopVarmus34•癌基因理论:每一个正常的细胞都带有能导致自己癌变的基因。在正常情况下,这些基因在控制细胞的增殖、分化中起重要作用,但一旦由于各种原因(如病毒感染,化学致癌物的作用等)导致这些基因发生突变的话,它们则可以引发细胞癌变。也就是说,细胞癌变的根本原因既不是病毒,也不是其他的致癌因素,而是存在于细胞自身的基因中。癌基因理论的确立是癌生物学研究中的一场伟大的革命。35病毒癌基因和对应原癌基因比较有序列的同源性和相似表达产物,但病毒癌基因经过病毒自身改变修饰,和对应的原癌基因比较,主要存在以下一些差别:363.原癌基因可分类生长因子类(如sis(PDGF-β))生长因子受体类(如erbB,trk)蛋白激酶类(如src)GTP结合蛋白类(如ras家族中的H-ras,K-ras)核蛋白类(如myc家族,fos家族,Jun家族)功能未知类根据蛋白的功能3738(三)原癌基因的表达调控•原癌基因在正常细胞中通常以单拷贝形式存在,正常情况下低水平表达或根本不表达。重排扩增点突变缺失normal功能或表达异常甲基化39401、点突变41Ras基因的点突变及转化活性分析Ras基因编码p21癌蛋白;几乎在其第12位和第61位的任何突变都能将c-ras原癌基因转变成各种转化癌基因。42LTR是反转录病毒基因组两端的长末端重复序列(longterminalrepeat):含有强启动子序列,当LTR插入原癌基因启动子区域或邻近部位后,可从根本上改变基因的正常调控规律。2、LTR插入4344原癌基因之间原癌基因与非原癌基因之间染色体数目和结构异常3、基因重排45abl原癌基因通过选择性染色体重排转变成细胞癌基因细胞分裂失控464、缺失(Deletion)oncogene(-)cancer一些原癌基因5’上游存在负调控序列,该序列的缺失或突变,则丧失抑制癌基因表达的能力。Burkitt’sLymphomac-myc因负调控序列缺失而过度表达。47•使每个细胞中基因拷贝数增加,从而直接增加可用的转录模板。•在肿瘤细胞中,DNA扩增事件的发生频率至少比正常细胞中高上千倍。•发生基因扩增的肿瘤细胞将获得选择性生长优势。5、基因扩增48TumorOncogeneAmplificationRcf.Small-celllungcancerc-mycUpto8OX67N-mycUpto50X66L-mycUpto20X266NeuroblastomasN-mycUpto250X295Glioblastomasc-crbB1(EGFR)Upto50X296Mammarycarcinomac-crbB2(HER2)Upto30X71.72c-mycUpto50X297CyclinD1Upto30X2016.原癌基因的甲基化改变DNA甲基化•DNA甲基化是一种表观遗传修饰,它是由DNA甲基转移酶(DNAmethyl-transferase,Dnmt)催化S-腺苷甲硫氨酸作为甲基供体,将胞嘧啶转变为5-甲基胞嘧啶(mC)的一种反应•在真核生物DNA中,5-甲基胞嘧啶是唯一存在的化学性修饰碱基•在人类基因组DNA中,约3%~4%的胞嘧啶碱基以5-甲基胞嘧啶形式存在•大约70%~80%的5-甲基胞嘧啶存在于CpG序列中,CpG二核苷酸集中的区域称之为CpG岛•限制性内切酶HapII和MspI均能识别CCGG,但不能识别CmCmGGDNA甲基化的生物学功能•通过基因启动子区及附近区域CpG岛胞嘧啶的甲基化可以在转录水平调节基因的表达,从而引起相应基因沉默,去甲基化又可恢复其表达•DNA的甲基化对维持染色体的结构和肿瘤的发生发展都起重要的作用,是细胞癌变过程中的重要一步肿瘤中DNA甲基化的作用•肿瘤局部相关基因的高甲基化:肿瘤抑制基因沉默•肿瘤中整体的低甲基化:肿瘤中整体的低甲基化•肿瘤组织相对于正常组织整体呈现低甲基化状态•目前有3种机制解释基因组整体的低甲基化在肿瘤中所扮演的作用:1.低甲基化导致原癌基因的去甲基化失活,导致癌基因的大量表达(Myc基因)2.整体的低甲基化是细胞染色体不稳定的易感因素3.低甲基化使肿瘤转移增加,DNA甲基化水平愈低肿瘤的浸润能力就越高,临床分期也愈晚肿瘤局部相关基因的高甲基化•近几年研究发现,在癌细胞中,肿瘤抑制基因甲基化的发生率与肿瘤抑制基因突变或缺失发生的概率大致相等,均可导致肿瘤抑制基因功能的丧失,如结肠癌、胃癌中hMLH1基因高甲基化导致DNA错配修复的缺失;脑肿瘤、结肠癌中MGMT基因的高甲基化导致化疗敏感性降低。•一些DNA甲基化抑制剂,例如5-氮杂胞苷和5-氮杂脱氧胞苷可以使失活的肿瘤抑制基因恢复其正常功能,为白血病和其他相关疾病的治疗提供了新的手段。肿瘤中DNA甲基化的应用•肿瘤筛查和风险评估:甲基化检测能够有效预测不同肿瘤中个体的风险率•肿瘤病理分型和治疗:甲基化指标的鉴定有助于肿瘤的临床病理分型,体外应用DNA甲基化抑制剂活化沉默的高甲基化基因•肿瘤治疗监测:一个或者多个
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码








 531244854
531244854
本文标题:肿瘤分子生物学
链接地址:https://www.777doc.com/doc-4760903 .html