您好,欢迎访问三七文档
当前位置:首页 > 建筑/环境 > 园林工程 > 第八章--植物营养遗传特性与改良
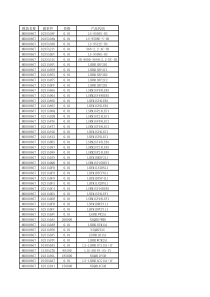
第八章植物营养遗传特性与改良进入21世纪,人口与生态环境的矛盾日益突出。随着人口的不断增长,人均耕地将越来越少,要在有限的耕地上生产出足够养活众多人口的粮食,必须提高作物单产。如果单纯依靠增加生产投入,既要消耗大量的人力和财力,又导致经济效率下降,甚至造成环境污染。充分发掘和利用植物自身的抗逆能力,通过遗传和育种的手段对植物加以改良,在有限的养分水分等资源条件下提高作物产量,提高资源利用率,是解决这一问题的有效途径之一。对于铁、磷等养分而言,土壤中的全量并不低,但常处于难以利用的状态,施人的磷肥和铁肥也易被土壤固定,利用率较低,利用养分高效植物,可以提高这类养分资源的利用率,减少投入。此外,世界范围内土地退化问题日趋严重,挖掘植物耐养分胁迫及毒害元素的能力,对于生态重建具有重要意义。在此背景下,植物营养遗传学应运而生并迅速发展。本章将着重介绍植物营养性状的遗传差异、生理基础和改良途径。在农业生产的范畴内,提高作物产量的基本途径有两种:①改良作物品种以适应它们所生长的环境条件;②改良环境以创造作物生长的最适条件。上述两种途径是相互补充、共同作用的。据估计,在60年代至80年代的20年间,一些发达国家的主要作物产量提高了2—3倍,增产的原因约有60%是由于品种改良,其余约40%是由于环境条件和栽培技术的改进,包括土壤改良和增施化学肥料等。无论是从经济效益还是从生态效益的角度来看,应用前一种途径提高作物产量比后一种途径更为有利。因为相对而言,环境条件或栽培技术的改良需要投入更多的能源、劳力和化学物质(例如化学肥料和农药),这不仅会使生产成本上升,而且还可能造成生态环境的破坏。因此,有人认为,我们今后的努力方向应是调整作物育种与耕作技术在提高作物产量方面的比重,将现有的比例(约60:40)提高至70:30甚至更高。所以,通过改良作物营养性状来提高作物产量应是农业科学工作者今后努力的一个方向,调整作物育种与耕作技术比重的潜力很可能就在植物营养性状的改良之中有幸的是,越来越多的人意识到植物营养性状改良的重要性,并已开展了一些颇有意义的工作。这方面的工作在我国也开始受到重视。在上述背景下,兴起了植物营养遗传学,近年来已有很大的发展。现代的植物营养遗传学研究的主要内容包括:①植物营养性状的形态和生理生化的研究。②植物营养性状的遗传潜力研究。③植物营养性状的遗传规律研究。④植物营养性状控制基因的染色体定位研究。⑤植物营养性状的遗传学改良研究。第一节植物营养的遗传变异性和基因潜力一、植物营养性状的表现型、基因型和基因型差异基因(gene)是控制生物生长发育性状的基本功能单位。它既是染色体的一个特定区段,又是DNA的一段特定碱基序列。碱基序列决定了植物的遗传密码。不同基因的差异在于其DNA碱基序列的排列组合不同,因而编码不同的蛋白质,行使不同的功能,进而控制生物体内的特定生理生化功能,在外观表现出不同的性状。不同生物个体内的基因组成不同,所有基因的组合称为基因型(genotype)。由于鉴定所有基因有实际困难,所以一般将生物体内某一性状的遗传基础总和称为基因型。在一定的环境条件下,基因表达使生物表现出某种性状,称为表现型(phenotype)。性状可由单基因或多基因控制。单基因控制的性状称为质量性状,而多基因控制的性状则为数量性状。农艺性状,包括很多植物营养性状(如耐低氮、耐低磷等),一般涉及植物体内的多个生理过程,受多基因控制,因而多数表现为数量性状。对于某一具体的生物学过程而言,如根细胞膜上的某种特定运输蛋白,则可由单基因或少数几个基因控制。对于某一具体性状,通常不能直接观察到其基因型,而只能通过观测表现型(可以直接观测到)来加以推测。因此,判断不同个体之间是否存在着基因型差异时,不能只是比较表现型,而必须推测确定各个体的基因型后才进行比较。对于质量性状(不连续变异的性状),可以通过表现型的分离和重组规律来确定其基因型。对于数量性状(连续变异的性状),由于一般是由多基因控制的,所以其基因型的确定比较困难,往往只能通过一些间接的方法来估测多基因综合作用的结果。这种综合作用的结果反映在某一性状的遗传变异程度上。一个性状由父代传递到子代的能力,可由遗传率(或称遗传力)来估测。一个性状的变异是由遗传变异和环境变异共同决定的。遗传率是指遗传变异程度在总变异程度中所占的比例,其中遗传变异程度是以遗传方差(基因型方差)来衡量,总变异程度包括了遗传方差和环境方差。二、植物营养性状的基因型差异例证在自然进化及人工选择过程中,由于分离、重组和突变等原因,某一生物群体的不同个体间在基因组成上会产生差异。群体中个体间因基因组成差异而导致的表现型差异通常被称之为“基因型差异”。由于表现型受环境影响很大,不同的基因型既可表现为不同的表现型。也可表现为相同的表现型。所以严格来讲,判断不同个体之间是否存在基因型差异时,不能仅限于其表现型的比较,而必须通过遗传学手段确定每一个体的基因型。植物在营养性状方面的基因型差异很早以前就引起人们的注意。1943年,Weiss发现生长在石灰性土壤上的有些大豆品系易出现典型的失绿症,而另一些则无失绿症状。Pope和Munger(1953)发现,芹菜对缺镁和缺硼的敏感性也存在着基因型差异。Graham在澳大利亚缺锌土壤上研究表明,小麦品种Aroona在缺锌时能取得较高的子粒产量,其锌营养效率高达92%;而品种Durati则不然,缺锌时产量仅为不缺锌时的41%(表10—1)。表10—1不同基因型小麦在缺锌和施锌条件下子粒产量比较品种供锌状况/t.hm-2锌效率﹡/%供锌不供锌Aroona1.421.3192Durati1.120.4541﹡锌效率(%)=缺锌处理的产量/施锌处理的产量×100在不同植物种类和不同品种之间,植物对土壤中铜利用效率有明显的差异(表10—2)。例如,小麦一般对缺铜都较敏感,而黑麦则对缺铜有较强的抗性。缺铜时,小麦品种Gabo和ChineseSpring产量极低,甚至颗粒无收,而黑麦的产量却相当高。小麦与黑麦的杂交后代小黑麦也表现出较高的铜营养效率,说明黑麦中存在高效吸收利用铜的基因,并能转移给小黑麦。表10—2在缺铜土壤上不同基因型对铜的反应植物种类品种每盆施铜量/mgb小麦00.10.440Gabo009.5100Halberb1.67.152.0100Chinesespring025.544.0100黑麦Imperial100114114100小黑麦Beagle98.695.293.6100数据为相对子粒产量;b每盆11kg。在表述不同植物利用养分能力的差异时常用养分效率(nutrientefficiency)这一概念,但对养分效率的具体内涵,则不同研究者尚无统一的定义。在同一物种不同品种之间,一般认为养分效率是指某个基因型在一定的养分供给条件下,获得高产的能力,或者认为,在同样的产量水平下,某个基因型需要的养分供应水平较低;而在不同物种之间,养分效率指当植物生长的介质中的养分不能满足一般植物正常生长发育的需要时(如土壤中养分元素的有效性较低),某一高效基因型植物能正常生长的能力。后一概念在生态学研究中更有意义。养分效率可以进一步划分为养分吸收效率和植物体内养分的利用效率。前者指植物吸收养分占养分总供应量的比率,后期则指植物体内单位养分所生产的干物质量或生物量,也称养分的生理利用效率。需要注意的是,植物体内养分的利用效率是一个复杂的动态的过程,涉及养分在植物体内的运输、分配、利用及再利用等多个生理过程,常常需要从全生育期的观点去分析,尤其是对于以收获繁殖器官为目标的植物。植物的营养性状基因型差异不仅表现在对养分缺乏的反应不同,而且还表现在对各种毒害元素的忍耐性的差异,如在酸性土壤中的铝、锰毒,在矿区及受污泥、污水污染的土壤中的锌、铜、硼、镉、镍等元素毒害,在盐碱地中的钠毒等。利用对重金属忍耐性强的植物,一方面可以恢复受污染土壤的植被,另一方面,还可能通过植物对重金属的富集,逐步地降低其在土壤中的浓度,最终达到土壤修复的目的,这称为“植物修复”(phytoremediation)。如遏蓝菜的一个种T.caerulescens即是一种重金属超累积植物,它可以在4000mg·kg—1含锌介质中生长,并在体内累积高达40000μg·g-1(干重)的锌,而与之同属的另一个种T.arvense在含锌500mg·kg-1的介质中就不能生长。第二节植物营养遗传的形态、生理学特性一、形态学和生理学特性植物养分效率的形成是一个复杂的过程,主要涉及植物对土壤中难溶性养分的活化、对土壤养分的高效吸收及植物体内养分的高效利用等。在实际研究中,不同的养分高效基因型的生理机制可能不同,如有的高效基因型吸收效率高,有的则利用效率高。而且即使都具有较高的吸收效率,有的基因型可能由于根量大,而另外一些则是因为根系分布合理。图10-2概括了养分效率基因型差异的可能机理。(一)吸收效率养分吸收效率既取决于根际养分供应能力及养分的有效性,同时也取决于植物根细胞对养分的选择性吸收和转运能力。在养分的吸收过程中,养分必须首先到达根表,才有可能被根吸收,养分离根越近,它被吸收的可能性越大。某些养分(如磷、锌等)在土壤中的迁移速率缓慢、迁移距离短,因此,植物根系的形态特征,如根系的体积、几何形状、长度、分布深度、分枝状况、根毛数量等和根系的结构特征对这些养分的吸收都有明显的影响。此外,许多植物还可以与真菌共生(如形成菌根共生体),通过菌丝的延伸,对土壤中有效性低的养分进行活化、吸收和运输。图10-2养分效率基因型差异的可能机理在养分供应不足的条件下,植物还可能产生一系列生理学特性的变化来改变根际的物理、化学和生物学性状,从而活化根际养分,增加根对养分的吸收。根际pH值和氧还电位的改变、根分泌的还原性和螯合性物质以及微生物能源物质的种类和数量等都能决定不同基因型植物对养分的吸收效率。不同植物种类,甚至不同品种之间在活化和吸收养分方面都有显著的基因型差异,这些差异可以在植物根际动态变化特征上充分地表现出来。这也是进行高效营养基因型植物选育的基础。现以根分泌物为例,说明不同基因型植物对营养胁迫的适应性及其调控机理。根分泌物是植物适应其生态环境的主要物质(表2),依据诱导因子的专一性,可把它划分为非专一性和专一性两类。1.非专一性根分泌物通过根系进入根际的非专一性分泌物可占植物光合作用同化碳的5%一25%。这些物质包括碳水化合物、有机酸、氨基酸和酚类化合物等,其分泌量受许多植物体内部和外部条件的影响,如能量、营养、通气状况、机械阻力、根际pH值和微生物的数量等。就养分状况来讲,缺乏磷、钾、铁、锌、铜和锰等都可能影响植物体内某些代谢过程,使低分子量的有机化合物(如有机酸、氨基酸等)累积并由根系分泌到根际。例如,缺磷使油菜分泌柠檬酸量增加。植物缺锌时,根细胞内铜锌超氧化物歧化酶(CuZn—SOD)的活性下降,而NADPH—氧化酶活性增加,致使细胞内氧自由基大量累积并产生毒害作用。过量氧自由基使细胞膜脂质产生过氧化作用,使膜结构破坏、透性增加,从而使根溢泌的低分子量的有机物如氨基酸、碳水化合物和酚类的数量和对铁、锰的活化能力都明显增加。这是非适应性机理中非专一性根分泌物在植物营养方面的典型例子。2.部分植物根分泌的低分子量有机物及其对根际养分的活化作用养分种类分泌物被活化的养分-P柠檬酸(羽扇豆)P、Fe、Zn、Cu、Mn、Al、Ca番石榴酸(木豆)P、Fe、Al-Fe酚类化合物(双子叶和非禾本科单子叶植物)Fe、Mn植物铁载体(禾本科植物)Fe、Zn、Cu、Mn、Al、PAl毒柠檬酸或苹果酸P2.专一性根分泌物专一性根分泌物是特定的植物受某一养分胁迫专一性诱导,在体内合成,并通过主动分泌进入根际的代谢产物。它的合成和分泌只受这种养分胁迫因子的诱导和控制,改善这一营养状况就能抑制或终止这种化合物的合成和分泌。当植物缺乏某一养分时,植物体可通过自身的调节能力,合成专一性物质并由根分泌到根际土壤中,促进这一养分的活化,以提高植物吸收利用效率,从而克服或缓解这种养分的
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码








 ckx
ckx
本文标题:第八章--植物营养遗传特性与改良
链接地址:https://www.777doc.com/doc-6093481 .html