您好,欢迎访问三七文档
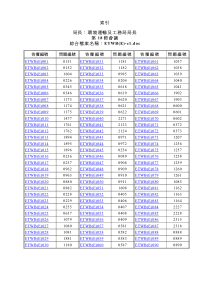
植物营养分子生物学及信号转导第一节植物无机氮转运系统和缺氮诱导的信号转导第二节植物应答低磷胁迫的机制及信号转导第三节植物钾营养吸收转运机制及信号转导第四节植物对铁元素的吸收运转调控及信号转导第五节植物镁元素营养的吸收转运机制与信号转导第六节锌铜锰分子生理学及基因表达调控印莉萍黄勤妮吴平主编《植物营养分子生物学及信号转导》,科学出版社,2006SulfurandAbioticStressTolerance植物硫代谢与抗逆性硫酸盐是土壤中硫营养的主要存在形式.在大气污染严重的地区,SO2与H2S可以进入叶片,但这只是硫营养吸收的次要形式。SO42-经过植物一系列还原与同化反应后进入有机骨架,生成半胱氨酸(Cys)。植物以Cys为前体,合成众多具有重要生物学功能的代谢产物,直接关系到植物耐逆境胁迫和农作物的产量与品质.硫Sulfur(S)生理作用1)蛋白质和生物膜的成分原生质的构成元素。2)酶与生活活性物质的成分辅酶A和硫胺素、生物素等维生素含有硫,也是硫氧还蛋白、铁硫蛋白与固氮酶的组分。3)构成体内还原体系蛋白质中-SH基与-S-S-互相转变,调节植物体内的氧化还原反应,还具有稳定蛋白质空间结构的作用。SulfurandSeedYield/Quality+S-SMycosphaerellagraminicola(majorpathogeninUSA)andPhaeosphaerianodorum(majorpathogeninEurope)SulfuranddiseasesWheatarchivelinksplantpathogenstosulphurdioxideBearchellS.J.,FraaijeB.A.,ShawM.W.&FittB.D.(2005).PNAS,102.5438–5442.Glutathioneplaysimportantrolesindiseaseresistance(PlantJ.49:159-172.2007)Glucosinolatemetabolitesasdiseasesignalingmolecules(Science323:95-101;101-106.2009)125mMNaCl(1week)100µMSulfate(2weeks)0µMSulfate100µMSulfateSulfurandsalttolerance“Sulfur-inducedresistance”硫Sulfur(S)缺硫症状植株矮小,硫不易移动,幼叶先表现症状,新叶均衡失绿,呈黄白色并易脱落。大豆植株矮小,新叶均一失绿发黄缺硫豇豆新叶失绿发黄油菜开花结实延迟玉米新叶失绿发黄缺硫1植物硫营养吸收运转和同化代谢2硫营养代谢在植物耐逆境胁迫中的重要功能3硫营养代谢的调控4硫营养信号转导植物硫代谢与抗逆性1植物硫营养吸收运转和同化代谢1.1硫酸盐的吸收运转和同化代谢1.2硫营养代谢中的重要代谢旁路及其功能1.3硫在植物体内的积累、贮存和再分配Sulfateuptake,transport,andassimilation1.1硫酸盐的吸收运转和同化代谢植物体内硫酸盐的同化可分为以下4步:(1)硫酸盐的吸收和转运;(2)硫酸盐的活化;(3)硫酸盐的还原;(4)Cys的合成。第4步是还原态硫进入代谢的唯一途径,从SO42-到S2-硫化物的还原仅发生在植物细胞的质体中。1.1.1硫酸盐的吸收和转运在根部细胞膜上的硫酸盐转运子的作用下,SO42-和H+按照1:3的比例以同向协同运输方式被主动吸收,质膜上的ATPase将细胞内H+泵出,由此形成的浓度梯度为硫酸盐的泵入提供动力;如果体外pH值升高,硫的吸收就被抑制模式植物拟南芥的硫酸盐转运子家族12个成员。根据编码序列的同源性、组织或细胞定位的不同以及动力学特性,它可分为4个亚家族:AtSultr1(Sultr1;1、Sultr1;2、Sultr1;3)、AtSultr2(Sultr2;1、Sultr2;2)、AtSultr3(Sultr3;1、Sultr3;2、Sultr3;3、Sultr3;4、Sultr3;5)AtSultr4(Sultr4;1、Sultr4;2)。AtSultr1-4分别负责不同组织和胞内硫酸盐的转运。拟南芥的硫酸盐转运子亚家族---AtSultr1AtSultr1亚家族的成员是高亲和力的硫转运子,主要但并非专一性地在根系表达。当环境中硫含量降低时,这个亚家族的成员在转录水平上受到调节。Sultr1;2和Sultr1;1有72.6%的同源性,负责拟南芥根部从环境中摄取硫酸盐与硫酸盐转运。Sultr1;2也可吸收转运硒酸盐。Sultr1;3参与拟南芥中硫酸盐的重新分布。它是特异表达于子叶和根系韧皮部的高亲和力硫酸盐转运子。低硫情况下,Sultr1;3在根系和叶片中的表达均增强。在Sultr1;3的T-DNA插入缺失突变体中,硫酸盐被阻滞在子叶中无法向地下部分转运,说明Sultr1;3负责硫酸盐从源到点的运输。拟南芥的硫酸盐转运子亚家族---AtSultr2AtSultr2亚家族的成员是低亲和力的硫转运子,主要表达于维管组织,负责硫元素的组织间分布。Sultr2;1和Sultr2;2这2个基因的表达模式在油菜和拟南芥中是有差异的。油菜中Sultr2;1在根、茎、叶中表达,而Sultr2;2在根系表达;拟南芥中二者均在根和叶中表达。AtSultr2;1表达于根木质部和中柱鞘以及叶片的木质部和韧皮部;AtSultr2;2在根的韧皮部和叶片的维管束中特异表达拟南芥的硫酸盐转运子亚家族---AtSultr3Sultr3;1、Sultr3;2、Sultr3;3和Sultr3;4转运子的具体功能不清楚。Sultr3;5与Sultr2;1共表达于根系维管组织,组成型表达的Sultr3;5本身的硫转运能力不明显,但是在低硫胁迫时,它可以辅助加强Sultr2;1的功能,促进SO42-由根系到地上部分的运输Sultr3;5的同源基因,它表达于固氮根瘤的共生膜上,主要用于对根瘤菌的硫运输和氮元素的固定。拟南芥的硫酸盐转运子亚家族---AtSultr4Sultr4;1在根系中组成型表达,不受低硫胁迫诱导;Sultr4;2受低硫诱导上调。Sultr4类硫转运子的主要功能是将液泡内的硫酸盐转运到胞质,在低硫条件下,又迅速将液泡贮存的硫酸盐释放到维管组织,从而调节根系组织液泡的硫贮存能力硫酸盐是如何进入叶绿体和质体几个硫转运子亚家族成员无一定位于叶绿体或质体膜上生物信息预测定位的准确性差,与实验结果误差很大。比如Sultr4亚家族被预测定位在叶绿体内膜上,但实验结果却显示定位于液泡膜上在叶绿体膜上仅发现Sultr1;1,而该转运子主要在根系中表达AkeyissueunresolvedSulfatetransportersinchloroplastenvelope?12sulfatetransporters(Sultr)inArabidopsisgenome:4subfamiliesSultr1:rootuptakeSultr2:vasculartransportSultr3:?Sultr4:vacuolarexchangeChloroplastenvelopeproteomerevealedonlyone:Sultr1.1Sultr1Sultr2(vascular)Sultr4(vacuolar)?Sultr3subfamily5SultrmembersLowaffinityandunabletocomplementyeastmutantLocalizationisnotknownExactroleinsulfatetransportisnotknownLocalizedinchloroplastsABCDEFLocalizedinchloroplastenvelopemembraneSULTR3;1-GFPGFPGFPChlorophyllMergeSULTR3;1-GFPWildtypeLoadingWesternblotSULTR3;1transportssulfateintochloroplastsSulfateuptakebypurifiedchloroplastsOthersulfatetransportersmustexistinchloroplast35SO4-2chloroplast35SO4-235SO4-2XSULTR3;11.1.2硫酸盐的活化进入植物体内的硫酸盐可被转运入叶绿体或质体,之后被活化而进入同化代谢。也可以进入液泡中贮存。植物体中硫酸盐的活化只需一步。在质体内,由ATP提供能量,ATPS(ATP硫酸化酶)催化SO42-生成APS(5'-腺苷磷酸硫酸)APS可以被APR(APS还原酶)还原成SO32-,也可以被APS激酶磷酸化为PAPS(3'-phosphoadenosine-5'-phosphosulfate)。拟南芥中APS基因家族拟南芥中有4个ATP硫酸化酶的编码基因:APS1(At3g22890)、APS2(At1g19220)、APS3(At4g14680)、APS4(At5g43780)。APS2(At1g19220)编码的是胞液ATP硫酸化酶,而其余3个ATPS均定位于质体,占叶片中ATPS酶总量的70%-95%。质体中的ATP硫酸化酶参与硫同化过程,胞液中的ATP硫酸化酶可能具有其它功能:如胞液中的硫酸盐化反应。1.1.3硫酸盐的还原--两步反应APS还原酶(APR)将电子从GSH转移到APS,进而生成SO32-在亚硫酸盐还原酶(SIR)作用下,SO32-从铁氧还蛋白(ferredoxin)接受电子产生S2-硫代磺酸硫代硫化物拟南芥中APR基因家族拟南芥中APR由3个基因编码:APR1(At4g04610)、APR2(At1g62180)和APR3(At4g21990)。植物体内的APS还原酶由2个结构域组成:N端的催化结构域及C端的电子传递结构域APR是硫酸盐还原途径的关键酶,植物体内APR的结构不同于硫酸盐还原细菌中APR的结构。植物体内的APR中不含有黄素,而是以[4Fe-4S]作为辅助因子。APR拟南芥中SIR基因拟南芥只含有1个定位于质体的SIR编码基因(AT5g04590),因而拟南芥中S2-主要是在质体部位生成。在光合作用组织中,亚硫酸盐还原酶(SIR)的电子供体是铁氧还蛋白。在非光合作用组织(根系)中,SIR的电子供体由NAD(P)H、ferredoxinNADP-还原酶以及ferredoxin协同提供FerredoxinNADP-还原酶将NADPH的电子转移到SIR,而特殊形式的铁氧还蛋白(Fd)和高的NADPH/NADP+比率是硫酸盐还原的关键。SIR1.1.4Cys的合成-*无机硫同化形成有机硫的关键步骤*硫-氮营养协调中的主要调控环节在serineacetyltransferases(SAT)和O-acetylserine(thiol)lyase(OASTL)的作用下,S2-和OAS生成Cys。OASCys合成酶(SAT/OASTL复合体)拟南芥中有5个编码SAT的基因:Serat1;1(At5g56760,SAT-c)、Serat2;1(At1g55920,SAT-p)、Serat2;2(At3g13110,SAT-m)、Serat3;1(At2g17640)和Serat3;2(At4g35640)。5个Serat基因主要在维管系统中表达(尤其是韧皮部)。Serat1;1、Serat2;1和Serat2;2在大部分组织中组成型表达。Serat1;1定位于胞液,Serat2;2定位于线粒体。Serat3;1和Serat3;2定位于胞液,且在多数组织中的表达都很微弱,在低硫和重金属胁迫下,Serat3;2的表达量变化显著,Serat3;1和Serat3;2主要用于胁迫条件下OAS的合成。在拟南芥中由3种OASTL基因编码OAS-TLA(At4g14880)、OAS-TLB(At2g4
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码








 41275601
41275601
本文标题:植物硫代谢与抗逆性
链接地址:https://www.777doc.com/doc-7363267 .html