您好,欢迎访问三七文档
当前位置:首页 > 医学/心理学 > 药学 > 韩春雨NgAgo论文中文翻译
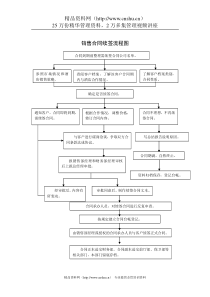
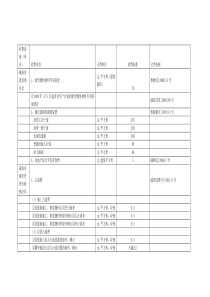
NgAgo——DNA导向的基因编辑技术摘要RNA导向核酸内切酶Cas9的出现使得基因组编辑成为一种广泛使用的技术。与此类似,来自于Argonaute蛋白家族的核苷酸内切酶也可以通过寡聚核苷酸的引导靶向降解入侵的遗传物质。我们在这篇文章中介绍了一种新的核算内切酶NgAgo,它被证明能够在人的细胞中进行基因的编辑。Ngago与5‘端磷酸化的24核苷酸构成的单链DNA结合,能够有效的在特定的位点形成基因组双链断裂。与Cas9不同的是,NgAgo-gDNA系统不需要PAM序列的参与,而且表现除了极高的特异性和在GC富集区序列极高的切割效率。RNA指导的核酸内切酶Cas9是最常用的基因组编辑工具,其通过gRNA与靶位点DNA的碱基互补配对的方式对靶位点进行精确的定位并切割靶位点。虽然Cas9系统进行了许多改进以提高其效率和特异性,但是脱靶现象和gRNA容易形成复杂的二级结构这些缺点还是影响了Cas9的发展。Argonautes是使用5'磷酸化的gDNA与靶位点结合并诱导DSBs形成的内切核酸酶家族,与Cas9类似,ago系统在基因表达的调控和对外来基因入侵的免疫过程中扮演着重要的角色。Ago与Cas9也有诸多不同的地方,首先,Cas9系统只存在于原核生物中,而Ago系统几乎存在所有生物中。虽然大多数的ago蛋白通过与ssRNA结合在RNA沉默中发挥作用。但是还有一些ago蛋白能够与ssDNA结合对靶位点进行一个靶向的切割形成DSBs。其次。在引导Cas9蛋白正确结合靶位点的过程中,gRNA需要在其3‘端形成一个RNA-RNA螺旋结构,而ago蛋白与靶位点的结合并不需要gDNA去形成复杂的二级结构。Cas9切割靶位点需要靶位点3‘端存在PAM序列。而Ago并无此要求。并且ago蛋白一旦与gDNA结合,它们会影响彼此的化学物理结构,作为一个整体要比比一般的核酸蛋白复合物表现出更为明显的动力学特征。基于这些特点,Argonautes系统可以形成多种工具蛋白,完成许多不同的任务。在这篇文章中我们介绍一种ago蛋白来自于N.gregoryi,(NgAgo)能够与gDNA结合,作为一种新的基因编辑工具在哺乳动物细胞中对基因组进行精确高效的编辑。结果NgAgo与guideDNA结合能够切割DNA由于大部分的ago家族的蛋白反映温度大于65度,限制了其在哺乳动物细胞中的应用,所以我们利用BLAST在数据库中找到了NgAgo蛋白。为了研究NgAgo蛋白是否与ssDNA结合,我们研究了从E.coli(同时产生磷酸化的ssDNA和ssRNA)中纯化出的NgAgo蛋白,核酸酶消化的结果证明了是DNA与NgAgo蛋白结合而不是RNA。(见图一a)测试由E.coli表达出的NgAgo蛋白是否能在体外37度的条件下切割靶位点DNA,我们设计了一个体外的酶切实验。我们设计了三个5‘端磷酸化的ssDNA,其中两个ssDNA能够碱基互补(FW与RV),并且对应于pACYCDuet-eGFP质粒上的位点,并且该质粒没有NgAgo的表达质粒上的同源序列(见图1b)。另外一个ssDNA(NC)是一段随机的序列并不对应于pACYCDuet-eGFP,我们还设计了一对5‘端磷酸化的ssRNA序列对应于FW与RV所对应的相同的位点。在酶切反应开始之前,我们将与NgAgo结合的天然的核酸代替为我们设计的guidessDNA。实验表明当不存在ssDNA或者与NC结合时,Ngago并不表现出酶切活性。当与FW和RV结合时,NgAgo能够线性化质粒。并且当ssDNA存在而ssRNA不存在时,NgAgo依然表现出了酶切活性。实验结果表明,NgAgo蛋白能够在与ssDNA结合后,在37度体外环境中对基因靶位点进行切割(如图1c)NgAgo蛋白与ssDNA结合能够在靶位点形成双链断裂为了评估NgAgo是否能够作为一种基因编辑工具在哺乳动物中发挥作用。我们第一次在293T中表达NgAgo蛋白并验证其是否能与内源性的核酸结合。在293T细胞中纯化出的NgAgo蛋白并没有发现核酸的结合。结果表明在人的细胞中5’端磷酸化的ssDNA的含量极少,表明内源性的ssDNA并不会导致NgAgo出现脱靶效应。然后,我们将5’端磷酸化的ssDNA和ssRNA转染到293T细胞中,并同时转染NgAgo的表达载体。研究表明,纯化得到的NgAgo蛋白只与5’端磷酸化的ssDNA结合。(见图2a)当我们在转染NgAgo表达载体24h后转染5’端磷酸化的ssDNA,NgAgo与ssDNA复合物的含量有了明显的减少,这表明NgAgo蛋白与ssDNA的结合发生在NgAgo蛋白表达后一个较短的时间内(如图二b)。并且,NgAgo并不能与ssDNA结合在体外37度的环境下,即便是从293T细胞中纯化出的NgAgo蛋白与ssDNA在37度孵育8h也不能导致NgAgo蛋白与ssDNA的结合(如图2c)。所以,NgAgo与ssDNA的结合只能在55度的体外环境下发生。并且只有在细胞中同时表达Ngago蛋白与ssDNA,形成的复合物能够对DNA进行切割,而从细胞中分离得到的NgAgo蛋白在体外与ssDNA结合形成的复合物并不能表现出酶切活性。此外,NgAgo蛋白与NCssDNA在体内结合后在体外与FW重新孵育8h也同样不能在靶位点诱导酶切的产生(如图2d)。这些结果表明NgAgo蛋白只会与第一次遇到的ssDNA结合,而不会发生ssDNA的替换在37度的条件下。这一点与哺乳动物自身的Ago蛋白家族类似。而NgAgo的这种特征就将其与非目标ssDNA结合的可能性最小化了。与重新装载ssDNA的NgAgo蛋白不同,在体内第一次装载ssDNA的NgAgo蛋白能够有效的线性化目标质粒载体。证明其能够在37度诱导DNA双链断裂的产生。而且我们认为55度的温度会使NgAgo丧失其酶切活性。为了确定NgAgo的酶切位点,我们将线性化的质粒载体加A尾后连接到T载体上并测序。结果表明。NgAgo蛋白在靶序列上形成了多个碱基的突变,而不是只打破了一个磷酸二酯键。这个结果与之前关于TrAgo的研究一致。去测试这种碱基突变是否基于NgAgo的核酸外切酶活性。我们延长了酶切反应的时间到72h,虽然在同样的条件下8h就可以完全的线性化质粒,结果表明目标序列没有发生降解。(图3a)测序结果表明,72小时的NgAgo反应时间后,在1~20个碱基的靶序列区域发生了碱基的随机突变。(见图3b)此外,NgAgo蛋白并不能切割线性化的质粒和DNA。这些结果表明NgAgo不是一种核酸外切酶。去探究是否NgAgo-gDNA系统能够作为一种有效的工具去对哺乳动物的DNA进行定点编辑,我们设计了一个体内的质粒酶切实验。我们共转染了pEGFP-N1质粒,NgAgo表达载体质粒和靶向于CMV启动子和eGFP编码区的ssDNA,我们通过评估eGFP的表达量来评估NgAgo的切割效率(见图3c)。为了将此系统与Cas9系统做对比,我们同时转染了pEGFP-N1和一个Cas9表达载体和sgRNa表达载体靶向于pEGFP-N1。通过对eGFP表达量的研究,我们认为NgAgo的效率与Cas9系统相差不大。同时利用这种方法我们得到了gDNA的最佳长度为24bp(见图3d)靶位点存在的广泛性与低的错配效应之后,我们探究了NgAgo系统在人基因组的内源性位点进行基因编辑的可能性。我们设计了一个改进的NgAgo蛋白,在其N端设计了一个核定位信号。我们设计了5gDNA对应于人类DYRK1A基因的第11个外显子。通过T7E1酶酶切去寻找内源性的基因突变。我们发现所有的gDNA都能高效诱导靶位点的切割的发生,同时我们通过测序证明了这一点。(见图4a)同时,我们针对8个基因设计了47个靶位点gDNA,所有gDNA的基因编辑的效率是相差不多的(21.3−41.3%;)(见图4b)。并且我们并没有发现NgAgo对于序列本身的性质有明显的偏好。我们同时在多种哺乳细胞中验证NgAgo-gDNA系统,NgAgo均可以在这些细胞的DYRK1A基因区诱发DSBs(见图4c)。为了研究NgAgo系统中,gDNA可能与靶位点存在的碱基错配的现象,我们在24nt的gDNA(靶向于DYRK1A基因)中的每个位置都引入一个单碱基突变。Ngago对于单碱基突变的gDNA特别敏感(切割效率降低73%~100%),并且等单碱基突变出现在8~11个碱基处时,切割效率降低85~100%。在gDNA中任何3个连续碱基的错配都将使NgAgo的酶切活性完全丧失(见图4d)。为了探究NgAgo对于单碱基的敏感度是不是会随着gDNA的长度的变化而改变,我们用同样的方法去研究了21nt的gDNA,单碱基突变在21ntgDNA的任何位置都会改变NgAgo的活性。并且第8到11个碱基同样的是最为敏感的区域,更为根本的影响NgAgo的活性。考虑到CRISPR/Cas9最多可以允许5个碱基的错配。这些数据,虽然只能初步的证明NgAgo所具有的低脱靶活性和高保真性。并且gDNA上第八个碱基的磷酸二酯键可能是NgAgo发挥核酸内切酶活性的重要部分,因为在第八个碱基上单碱基的突变就可以完全使NgAgo丧失核酸内切酶活性。我们利用哺乳动物中的DYRK1A基因来比较CRISPR/Cas9系统与NgAgo系统的切割效率,因为该基因已经在Cas9的研究中广泛应用。NgAgo系统和Cas9系统在该位点的切割效率相差不大。(Fig.4e,31.97%forNgAgovs.32.2%forCas9)(见图4e)然后,我们测试了NgAgo系统是否在GC富集区进行切割要比Cas9系统更有优势。因为gRNA在这些区域更容易产生高级结构,而DNA则不会,这就会影响gRNA与Cas9复合物的形成。结果表明在GC富集区,NgAgo的切割效果明显好于Cas9系统。这就证明了NgAgo在基因编辑区域的选择上面较Cas9系统更有优势。此外,根据相关的研究和我们的实验,Cas9系统发挥作用需要在靶位点的下游存在PAM序列,而NgAgo并无此要求。NgAgo在基因敲入中的应用当我们验证了Ngago能够在哺乳动物基因组中的靶位点上发生切割,我们下一步开始探究是否可以进行基因片段的敲入。在基因研究中,基因的定点敲入有着广泛的应用。我们设计了一个donorDNA片段上面有一个报告基因和一个G418抗性基因(见图5a)。报告基因有两个阅读框,一个编码mRFP一个编码eFGP,中间用一个TGA终止子隔开,这个报告基因区由于没有启动子的存在,所以它们不会表达除非将它们整合到一个哺乳动物的基因中,我们将靶位点设在了DYRK1A位点上,同时将NgAgo表达载体,gDNA和donorDNA转染到293T细胞中。我们发现细胞中有红荧光蛋白的表达。通过PCR和测序我们确定donorDNA整合到293T细胞的基因中(见图5b)。通过G418筛选,我们得到了敲入的阳性细胞。然后我们重新利用NgAgo系统靶向于TGA终止子区,这会导致TGA终止区出现非同源末端连接修复现象。使得mRFP和EGFP同时表达(见图5c)。后续的研究表明,共有11.7%的细胞能够表达绿荧光蛋白。这证明了NgAgo在哺乳动物基因编辑中的高效率。当我们同时在细胞中利用Cas9系统和NgAgo系统引入基因片段的敲入时,Southernblot的结果表明Cas9介导的基因敲入存在着非靶位点的敲入,NgAgo并没有出现这一现象。
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码








 310272
310272
本文标题:韩春雨NgAgo论文中文翻译
链接地址:https://www.777doc.com/doc-7514390 .html