您好,欢迎访问三七文档
当前位置:首页 > 行业资料 > 交通运输 > 植物逆境生理-第五章-光损害作用与植物的防护
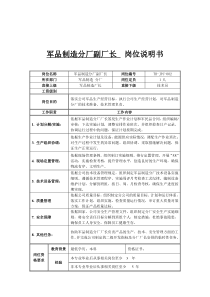
第五章光损害作用与植物的防护第一节太阳辐射对植物的损害第二节植物对光损害作用的防御及对局部辐射状况的适应第三节紫外线辐射对CHS基因表达的调节及信号传导第五章光损害作用与植物的防护第一节太阳辐射对植物的损害第一节太阳辐射对植物的损害一、太阳辐射与辐射光谱二、光抑制三、紫外线辐射的损害作用第一节太阳辐射对植物的损害一、太阳辐射与辐射光谱(一)太阳辐射太阳一刻也不停地向茫茫宇宙空间辐射着大量的电磁波。其中射向地球的那一部分,向地球输送了大量的光和热。据粗略估计,太阳每分钟向地球输送的热能大约是250亿亿卡,相当于燃烧4亿吨烟煤所产生的能量,这是非常可观的。地球在一年中从太阳获得的能量,相当于人类现有各种能源在同期内所提供的能量的上万倍。地球上的一些天然能源(如煤、石油等)可能有枯竭的那一天,而太阳能却是取之不尽,用之不竭的。太阳能是人类可以无限使用下去的一种巨大的天然能源。地球除了从太阳那里取得能源外,还可从其它天体,如月球等取得能量,但其数量是微不足道的。地球从月球等其它天体所得的辐射能,仅为太阳的亿分之一。而来自宇宙的辐射能也仅为太阳辐射能的20亿分之一;从地球内部传到每平方厘米地面上的热量,全年才为5.4卡,仅为来自太阳辐射能的万分之一。所以说,太阳是地球和大气能量的源泉。(二)太阳辐射光谱及光损害太阳是个炽热的大火球,它的表面温度可达6000°C,它以辐射的方式不断地把巨大的能量传送到地球上来,哺育着万物的生长。太阳辐射的波长范围,大约在0.15~4微米之间。在这段波长范围内,又可分为三个主要区域,即波长较短的紫外光区、波长较长的红外光区和介于二者之间的可见光区。太阳辐射的能量主要分布在可见光区和红外区,前者占太阳辐射总量的50%,后者占43%。紫外区只占能量的7%。在波长0.48微米的地方,太阳辐射的能力达到最高值,数值约为3.0卡/cm2以上。图5-1太阳辐射光谱组成在自然界,太阳辐射对植物的损害作用一般发生在两种情况下,一种情况是植物被较长时间地曝露在强度过高的可见光下,特别是在某些其它环境所造成的逆境条件下,所受到的伤害更为明显,即光抑制现象(photoinhibition);另一种情况是由太阳辐射中紫外线所引起的损害。近年来,全球正在出现臭氧层的逐渐消蚀,臭氧层的减少会导致更多的紫外线照射到地球表面上,因此紫外线辐射生态学正在引起人们的极大关注。二、光抑制(一)光抑制现象虽然光是植物光合作用的根本推动力,没有光植物便不能进行光合作用,光不足则不能高速地进行光合作用,可是光过剩对植物来说也不是好事。当叶片接受的光能超过它所能利用的量时,光可以引起光合活性的降低,这就是光合作用的光抑制现象。它的最明显特征是光合效率的降低。在没有其它环境胁迫的条件下,晴天中午许多植物冠层表面的叶片和静止的水体表层的藻类经常发生光抑制。由于发生光抑制的前提是光能过剩,所以,任何妨碍光合作用正常进行而引起光能过剩的因素,如低温、干旱等,都会使植物易于发生光抑制。因此,在两种或两种以上环境胁迫因素同时存在时,即使在远低于光合作用饱和光强的中低光强下植物也会发生光抑制。光抑制也不是用肉眼可以看到的。冬季一些常绿树木冠层表面叶片变黄变白是由于其中的叶绿素的氧化破坏,那是低温条件下叶片长期经受严重光抑制的结果。光氧化虽然不同于光抑制,但往往同光抑制有密切的联系。光抑制大体上有两种不同的情况:一种情况是,引起光抑制的胁迫因素去除后,光合功能在1h左右便可以完全恢复。这种光抑制主要是光合机构通过叶黄素循环或反应中心的可逆失活等过程耗散过剩光能的反映。在只有强光一种胁迫因素存在的自然条件下,植物的光抑制大多属于这种情况。另一种情况是,引起光抑制的胁迫因素去除后,光合功能往往经过许多小时甚至几天才能完全恢复。这种光抑制主要是光合机构受到破坏的结果。在这种情况下,不仅叶片的光合效率降低,而且光饱和的光合速率即光合能力也降低。恢复过程的长短取决于光抑制期间的光能过剩程度和持续时间的长短,以及恢复期间光照、温度和水分等条件是否合适。弱光比强光和黑暗条件有利于恢复。恢复的速度也因植物种类和生长条件不同而异。(一)光合机构的破坏1.PSII的破坏近些年的研究普遍认为,光破坏的原初作用位点在PSII反应中心,而QB蛋白的降解只是它的一个结果。目前研究从分子水平已取得一些重要进展。Durrant等(1990)认为PSII光破坏是由电荷分离形成的离子P680+与Pheo-发生电荷重组时产生1O2破坏造成的,1O2能够使PSII反应中心D1/D2/Cyt-b559复合物His残基及Pheo破坏。进一步研究表明1O2首先使Pheo分子破坏,然后使PSII原初电子供体P680破坏。在PSII反应中心的破坏中,D1和D2蛋白也发生了化学断裂和共价交联。此外,PSII膜复合物蛋白二级结构也被破坏,特别是螺旋构象伤害严重,而对折叠和转角影响不大,进一步研究指出PSII光破坏不只是蛋白和色素的破坏,而且也存在膜脂分子的伤害。以前,人们认为光破坏过程中D1蛋白的降解与PSII中存在的蛋白酶有关,但近来用丝氨酸类蛋白酶抑制剂二异丙基氟磷酸(DFP)并采用同位素标记的方法,研究表明光破坏过程中D1蛋白的降解可能还与核心天线CP43有关。2.PSI的破坏近年来,有人报道高等植物活体内存在PSI选择性抑制活性,即在低温条件下PSI对光更敏感,比PSⅡ更易发生光抑制。从此人们对PSI光抑制的相关研究才逐渐重视起来,认识到PSI光抑制的生理意义,尤其是对低温下PSI的光抑制更加重视。低温胁迫下,PSI是马铃薯叶片和黄瓜叶片电子传递的限速步骤,PSI发生光抑制时,两种植物都需O2的参与。Gerber和Burris的试验表明,强光处理硅藻Amphorasp.可引起光氧化态的P700水平下降,而且光合速率的下降(14CO2标记)大于氧化态P700的下降幅度,表明PSI的某些组分可能发生严重失活。Inoue等(1986)认为,有氧条件下,光抑制的位点是铁硫中心,而铁硫中心能限制NADP+的光还原;相反,在强还原条件下,随着NADP+光还原程度的下降,铁硫中心、维生素K(VK)和P700的水平则没有改变,表明此时光抑制位点可能位于A0和FX之间。活体PSI选择性光抑制只发生在低温下。当冷敏感植物如黄瓜的叶片暴露在100~300mol·m-2·s-1光强和低温下(0~10℃)时,光合速率下降与PSI光抑制对温度的依赖之间有很密切的相关性,因此推测叶片光合速率下降的主要原因是PSI光抑制。有试验认为,在PSI受害过程中,P700受体侧的Fe-S中心最先受到伤害,然后是P700本身,同时psaB基因产物也出现降解。psaB基因产物的降解可能与活性氧的产生有关。Baba等对菠菜叶片PSI颗粒水平的研究发现,强光(11000mol·m-2·s-1)下叶绿素发生漂白,反应中心蛋白降解,电子传递链活性降低。Tjus等(1998)的试验表明,PSI的A、B蛋白亚基的降解是在电子传递辅因子受到破坏之后发生的,即蛋白亚基的降解可能不是PSI光破坏的最初结果。据此,他们推测PSI光抑制破坏顺序应包括:1)末端电子受体FA/FB的伤害程度轻微,这从P700毫秒级衰减可以看出;2)伤害程度较大的是FX的破坏,因而呈微秒级衰减;3)可检测到的P700光吸收完全丧失,这可能是P700直接受破坏或因A1的破坏而导致由A0向P700的快速逆反应造成的。这进一步证实了Sonoike和Terashima的观点,即在黄瓜叶片内光抑制的主要作用位点可能是FX或A1。但是,与微秒级衰减相比,毫秒级衰减的相对破坏作用要小,这暗示一旦FA/FB被破坏,FX就会变得不稳定,并很快衰变。据此说明,FX可能是最初作用靶位,但破坏作用是由活性氧造成的,于是Tjus等推测FA或FB比FX在PSI反应中心的位置更暴露,因而更有可能是最初作用位点,且FB对化学失活很敏感。尽管已经有一些试验表明PSI在特定条件下会发生光抑制,但是这些PSI活性比PSII下降幅度大的试验结果似乎还不能充分说明PSI是低温作用的最初靶位。低温下PSI光抑制在光合作用下降过程中所占比例有多大?在自然条件下,PSI的光抑制是不是普遍存在?常温条件下的PSI光抑制是否也会先于PSII发生,如果常温条件下发生PSI光抑制,其生理意义是什么?胁迫条件下,两个光系统间的相互作用机制如何?等等,都有待于进一步研究。这些问题的阐明将有助于了解PSI光抑制的作用机制和胁迫条件下PSI和PSII之间的相互关系。三、紫外线辐射的损害作用太阳辐射是影响生物生长发育的主要因子。生物在从太阳辐射中获得光和温度的同时,不可避免地受到短波光,即紫外辐射的伤害。紫外辐射作为生物生长发育中重要的环境因子,它与生物之间有着密切的联系。研究生物与紫外辐射的相互关系的科学叫做紫外线辐射生态学。(一)大气臭氧层减薄与地表紫外辐射增强紫外线是不可见的非电离辐射,其波长范因为200~400nm。人眼对波长为380nm的辐射开始有光觉,而波长短于100nm的辐射实际上能电离所有分子。辐射靠光子输送能量,一个光子的能量E为:E=h=hc/式中是普朗克常数(6.6210-27erg·s),是频率,c是光速,是波长。光子的能量E和频率不随输送辐射的媒质而变,但光速c和波长是随之而变的。已知1eV(电子伏特)=1.610-12erg。相应于波长为1240nm的光子能量,若光子能量以电子伏特为单位,波长以纳米为单位,E=h=hc/可改写为:E=1240/由此式不难求,相应于波长为380~190nm的光子能量为3.1~6.2eV。紫外线的波长下限是100nm,但其生物学效应的波长下限为200nm,再短一些将为空气和水强烈吸收。而大多数有机分子要波长在150~180nm以下的紫外线才能使之电离,所以紫外线对生物物质是非电离辐射。地表紫外辐射能量占太阳总辐射能的3%~5%。紫外辐射的波长范围为200~400nm,根据其生物效应分为短波紫外辐射(又叫超强效应波)(UV-C,200~280nm),中波紫外辐射(又叫强效应波)(UV-B,280~320nm)和长波紫外辐射(又叫弱效应波)(UV-A,320~400nm)。UV-C对生物有强烈影响,但它在平流层中基本上被臭氧分子全部吸收而不能到达地面。UV-A可促进植物生长,一般情况下无杀伤作用,它很少被臭氧吸收。从生态学角度分析,UV-B是非常重要的,它对生物具有较强的伤害作用,在臭氧层正常时仅有10%可抵达地球表面,但随着臭氧层的破坏,其对地球表面生物的危害越来越大,从而产生了一门新的学科——辐射生态学。大气臭氧层能吸收强烈的太阳紫外辐射,使地球生物得以正常生长,从而成为地球生命的有效保护层。近些年来,由于人类大量使用氯氟烃(CFC)等一类致冷剂、发泡剂、清洁剂、喷雾推进剂等,使臭氧浓度降低,造成臭氧层破坏,甚至出现空洞。二十多年前,人们注意到平流层臭氧衰减,近十年全球臭氧层平均减少了2%~3%,并且还在加剧。臭氧层快速减薄已经通过卫星得到证实,最明显的减薄发生在南极大陆上空,在春季,衰减率高达71%。过去十年我国北京香河和昆明两个监测站的监测结果表明,臭氧总量呈降低趋势,分别降低了5%和3%。据估计,如不加任何控制,发展到2075年臭氧层将减少40%。一个严重的后果就是到达地表的紫外辐射显著增加。平流层臭氧衰减导致更强的具有生物学效应的UV-B辐射到达地表已被证实。在300nm处,臭氧层减薄将导致光谱辐射倍增。卫星资料表明,在1979~1993年,南北半球的中高纬度地区都有显著的UV-B辐射增加。根据1981~1989年在瑞士阿尔卑斯山的测量,UV-B辐射每年增加1%。80年代后期与1980年相比,东京的UV-B辐射增加了30%。在多伦多,1989~1993年间,300nm附近的辐射,在冬季每年增加35%,而夏季每年增加7%。我国北京1980~1989年及昆明1980~1990年的观察也表明UV-B辐射与臭氧含量有相反的变化趋势,总体上是臭氧含量降低,
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码









 wxu1989
wxu1989
本文标题:植物逆境生理-第五章-光损害作用与植物的防护
链接地址:https://www.777doc.com/doc-1850248 .html