您好,欢迎访问三七文档
当前位置:首页 > 商业/管理/HR > 公司方案 > 端粒酶与肿瘤的研究进展
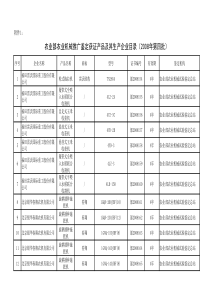
1端粒、端粒酶及其肿瘤关系的研究进展陈晨93K5B指导教师:于爱鸣【摘要】近些年来研究发现,端粒和端粒酶与人类的肿瘤的发生发展有着极为密切的关系。端粒是线性染色体末端的一段特殊的DNA-蛋白质复合物;端粒酶是一种逆转录酶,其能以自身RNA为模板合成端粒,它的存在解决了染色体末端复制缩短问题,同时认为端粒酶的过度表达与细胞的永生化和癌变直接相关。端粒酶抑制剂的研究和开发为肿瘤治疗提供了新的思路,并有可能成为肿瘤治疗的突破。本文主要介绍了端粒和端粒酶的结构和功能及其与肿瘤的关系,以及端粒酶抑制剂在肿瘤方面的应用。【关键词】端粒;端粒酶;肿瘤治疗;抑制剂【正文】长期以来,普遍认为癌症是由破坏细胞正常生长调节的多种基因突变引起。尽管不同的癌症可能涉及到一个特定的基因突变,个体的癌症往往表现为以多基因突变为特征。除了这种特殊的病因学外,最近的研究支持一种新的模式,即端粒酶的激活可能是许多恶性肿瘤发生的先决条件[1]。近年来端粒酶抑制剂的研究和开发为肿瘤治疗提供了新的思路,并有可能成为肿瘤治疗的突破。一、端粒和端粒酶(一)端粒的结构和功能早在上个世纪30年代,赫尔曼·马勒(HermannMuller,1946年诺贝尔奖获得者)和芭芭拉·麦克林托克(BarbaraMcClintock。1983年诺贝尔奖获得者)发现了染色体末端的一个特殊结构,他们认为该结构可以阻止染色体彼此黏附,并设想其可能具有保护性作用[2]。Muller将这种天然末端定义为端粒,她用希腊文的telos(末端)和meros(部分)创造了telomere(端粒)一词[3,6]。端粒是真核细胞染色体末端的DNA重复序列和特异结合蛋白的复合体。在真核生物中广泛存在。不同物种的端粒序列有所不同,但较相似。人类端粒DNA由基本序列单元-(TTAGGG)n-反复串联而成,总长度约为2-15Kb。由于富含G碱基单链的不完全复制以及富含C碱基单链的特殊反应,端粒在细胞的每一次分裂过程中都会失去几十个碱基序列。当端粒缩短到一定程度时,细胞就开始衰老或者步入死亡[12,13,14]。端粒DNA主要功能有:第一,保护染色体免受核酸酶降解;第二,防止染色体融合、降解、重组等损伤;第三,为端粒酶提供底物,解决DNA复制的末端隐缩,确保染色体的完全复制。(二)端粒酶的结构和功能端粒酶是1987年由Greider和Blackburn在四膜虫细胞核提取物中首先发现并纯化的。人的端粒酶是由Morin于1989年首次在人宫颈癌Hela细胞中发现并鉴定的。目前,已分离的人的端粒酶包括3个主要组分:端粒酶RNA(humantelomeraseRNA,hTR)、端粒酶相关蛋白(Telomerase-associatedprotein,TEPl)和端粒酶反转录酶(humantelomerasereversetranscriptase,hTERT)。人的端粒酶RNA(hTR)序列在1995年被克隆,发现它为单拷贝基因,由450个核苷酸组成,模板序列为5'-CUAACCCUAAC,其与人细胞端粒的反复串联基本碱基序列单元-(TTAGGG)n-互补,从而能够以自身为模板催化指导端粒DNA的合成,解决染色体的末端复制问题,防止端粒的缩短。二、端粒酶与肿瘤(一)端粒酶与肿瘤的发生——端粒、端粒酶假说早在l970年,苏联科学家就曾经提出:染色体末端逐渐丢失,细胞很可能会因染色体的缩短而最终退出细胞周期[4]。在肿瘤发生的早期,端粒长度大幅度地缩短,而端粒的缩短又反过来影响肿瘤的发生[5]。研究表明,端粒酶的激活可能是体细胞向肿瘤细胞转化的关键步骤,是细胞癌变的早期事件。近年来,端粒、端粒酶假说受到了广泛的关注和支持。大部分正常体细胞不表达端粒酶活性,随细胞分裂次数增加,端粒进行性缩短。端粒的缩短也许是生物体细胞有丝分裂阶段的一种调节措施,其中端粒酶起了主要的调控作用。由于染色体在每次细胞分裂中是不完全复制的,染色体末端序列不可避免地会在某一点上发生进行性丢失,这个位点不再保护染色体的末端免受重组或降解。1990年Harley提出端粒、端粒酶假说[15],该假说认为:端粒于每次细胞的有丝分裂过程中的进行性丢失是正常的,端粒的长度作为一个决定细胞剩余复制能力的钟(clock)。端粒缩短至一定程度时,复制钟停止运转,细胞停止增殖而老化[7,8]。这可能是由于端粒的丢失激活p53及其下游基因如p21和Rb诱导细胞周期停滞或凋亡的缘故,称为第一死亡率2期(firstmortalitystage,M1期)。现已证实,若发生p53或Rb突变、SV40的T抗原突变,则可使细胞绕过(bypass)第一死亡率阶段而继续分裂(约50次),此时端粒酶仍为阴性,端粒仍在进一步缩短。之后到达第二死亡率期(secondmortalitystage,M2期),或称“危象”(crisis)[4,8]。此时大部分细胞老化或死亡。极少的细胞能超越M2状态获得永生,它们通常表达端粒酶的活性。正常人性细胞和少数处于高增殖状态的体细胞,包括皮肤基底细胞[9],造血系统干细胞和成熟血细胞都具有端粒酶活性,但只有性细胞表达足够的端粒酶活性,能完全维持端粒长度[10,11]。可见,端粒酶可能为细胞永生化和大多数恶性肿瘤增殖所必需。事实上,约80%~90%的人恶性肿瘤细胞中检出端粒酶活性,提示端粒酶不仅为一种新的癌标记物.而且成为癌症治疗的新靶点[7]。这一假说已被越来越多的事实所证实:Harley等发现人的成纤维细胞的端粒长度随着年龄的增长而不断地缩短[16]。Hiraga等发现只有具有端粒酶活性的神经上皮瘤者可以建立细胞株,而没有端粒酶活性的神经上皮瘤则不能建立细胞株[17]。潘立峰等在乳腺癌组织中检测端粒酶表达阳性率为91.7%,而且与肿瘤的大小,淋巴结的状态,临床分期有相关性[18]。因此,端粒酶重新激活是癌变细胞一个带有普遍性意义的生物学标志,被认为是细胞癌变的一个重要细胞生物学异常事件,是细胞永生化及肿瘤发生的关键步骤[19,20]。(二)端粒酶作为肿瘤标志物早期诊断肿瘤现今,端粒、端粒酶假说已经得到了较多的认可,从一个全新的方面解释了肿瘤的发生和发展。研究发现,人恶性肿瘤细胞中染色体的端粒和端粒酶的活性均不同于除一些更新组织的增殖细胞外的正常体细胞。在许多正常体细胞中检测不到端粒酶活性或活性极低,而几乎所有的人类恶性肿瘤细胞中的端粒酶均呈现活性。人端粒酶催化亚基mRNA在大多数正常体细胞中不表达,但在原发性肿瘤、癌细胞系中却有强表达[21,22]。所以端粒酶可以作为各种恶性肿瘤细胞的一个共同的分子标志物[23]。很多学者认为通过相关组织端粒酶活性的检测有助于肿瘤的诊断。Christopher等对甲状腺细针抽吸结果不确定而进行甲状腺切除术的100个病人进行hTERT测定,发现48例甲状腺癌病人hTERT阳性率90%,52例良性病变hTERT阳性率为35%。当把10例并发淋巴细胞性甲状腺炎的病人除外,hTERT的敏感度为91%,特异度为79%。仅根据hTERT的结果确定手术方法,可减少甲状腺叶切除术7例及全甲状腺切除术6例[24]。因此,术前hTERT测定对确定甲状腺肿瘤的良恶性有一定的诊断意义。何奇应用改良的端粒末端重复序列扩增(telomericrepeatamplificationprotocol,TRAP)银染法,检测端粒酶活性的结果发现,39例大肠癌阳性率为89.7%,21例溃疡性结肠炎阳性率为9.5%,22例正常对照组阳性率为0%。因此,端粒酶可作为理想的肿瘤标志物在大肠癌的早期诊断中具有重要作用[25]。李玲、马俊芬等使用TRAP方法对骨肿瘤的组织标本进行端粒酶活性的检测。62例骨肿瘤冻存标本中,端粒酶活性有48例阳性(79.0%)。同时检测了19例的肿瘤旁组织,未发现阳性。3例骨软骨瘤有l例阳性[26]。该结果表明端粒酶活性可作为涎腺恶性肿瘤早期诊断的分子指标。岳岩等利用TRAP检测了83例眼眶恶性肿瘤组织及癌旁组织、165例眼眶良性肿瘤组织和6例正常眼眶组织的端粒酶活性。结果显示,254例(337份)标本中,眼眶恶性肿瘤组织、癌旁组织、良性肿瘤组织的端粒酶活性及hTERTmRNA阳性率分别为95.2%(79/83)、97.6%(81/83);3.6%(3/83)、4.8%(4/83);3%(5/165)、3.63%(6/165);6份正常眼眶组织未检测到端粒酶活性及hTERTmRNA的表达。说明眼眶病变组织端粒酶活性及hTERTmRNA表达水平的检测对眼眶肿瘤早期诊断具有应用价值[29]。此外,在胰腺癌、肺癌、食道癌中也得到相似结果,认为hTERT是一个较理想的诊断指标。(三)端粒酶与肿瘤的治疗由于端粒酶在肿瘤细胞中的高表达,在正常体细胞中的不表达或极少表达,现已逐渐得到人们的重视,成为了当今肿瘤治疗的一个新靶点。目前基于端粒、端粒酶假说以及对端粒酶结构、功能的理解,认为抑制端粒酶活性或基因表达的抑制剂,使肿瘤细胞不能有效合成端粒序列,从而发生“凋亡”,达到治愈肿瘤的目的[27]。现今主要有两种途径:以端粒酶RNA为靶点;以端粒酶hTERT为靶点。1.以端粒酶RNA为靶点(1)应用反义寡脱氧核苷酸(Antisenseoligodeoxynucleotides)[20,27]端粒酶RNA序列中含有与DNA互补的模板序列,因此可设计能与之结合的反义寡脱氧核苷酸来使端粒酶失活,从而阻止端粒序列的合成。反义寡脱氧核苷酸包含与目标RNA互补的片断,其竞争性与目标RNA相结合,一旦两者互补杂交就会激活RNA酶H(RNaseH)来降解杂交体。设计有针对端粒酶的RNA的模板或非模板区,因为端粒酶在合3成端粒时,其RNA成分要暴露出来,这就为反义寡脱氧核苷酸与其结合形成杂交体提供可能。应用这种方法最早在HeLa细胞中成功地抑制端粒酶的活性[28]。Mukai等设计出通过反义寡脱氧核苷酸结合RNA摸板区域以阻断端粒酶活性的方法,将5-磷酸化2~5寡腺苷对裸鼠的颅内胶质瘤进行处理,发现恶性胶质瘤细胞的增殖能力降低至43%-76%[30]。王红军等应用针对端粒酶的RNA反义寡脱氧核苷酸,24小时后的凋亡指数为(23.01±2.98)%[32],与对照组相比显示出明显的治疗效果。(2)锤头状核酶(teloRZ)锤头状核酶是一种存在于细胞核具有核酸内切酶活性的小分子RNA,它由一个反义序列和位于其旁边的催化核心构成。其反义序列不仅能识别目标RNA的位点与其结合,而且特异性的将其水解,又称“基因剪刀”。Yokoyama引等针对hTR3'端不同序列为靶点设计了3个锤头型核酶,将其RNA导入Ishikawa子宫内膜癌细胞,细胞的端粒酶活性降低,其中以模板区为靶点的核酶具有最强的降解端粒酶活性的效力。并导致了端粒的缩短[31]。核酶与一般反义寡核甘酸相比较,抑制端粒酶活性的作用较为确切,有望成为新一类的抗癌药。2.以端粒酶催化亚基hTERT为靶点的免疫治疗在端粒酶亚基中,hTERT是决定端粒酶活性的关键因素,与肿瘤的关系更为密切。因此,以hTERT为靶点的肿瘤新的治疗手段越来越受到重视。早在1999年,Vonderheide等首先发现来源于hTERT的9肽与MHC-I类分子结合递呈在肿瘤细胞表面,并作为抗原特异性激活细胞毒性T细胞(CTL)导致肿瘤细胞溶解,因而初步认为,hTERT是一种可被CTL特异识别的广泛表达的肿瘤相关抗原[33],而开创了以端粒酶为靶点的肿瘤免疫治疗新方法。郑骏年等以针对hTERT基因的小干扰RNA转染肾透明细胞癌株786-0细胞,发现hTERT基因mRNA、蛋白表达水平均明显受抑制,同时肿瘤细胞增殖抑制、凋亡增加[34]。同样,来源于端粒酶hTERT,HLA-B0702限制性的多肽能够诱导人CTL反应,且hTERT多肽特异性CD8+CTL能杀伤多种hTERT阳性的肿瘤细胞,应用hTERT,HLA-B0702限制性的多肽能诱导老鼠体内CTL的免疫反应[35]。Filaci等研究表明,以端粒酶为靶基因治疗的癌症病人,大多数外周都出现CD8+T细胞
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码









 2mhwdjru
2mhwdjru
本文标题:端粒酶与肿瘤的研究进展
链接地址:https://www.777doc.com/doc-2240419 .html