您好,欢迎访问三七文档
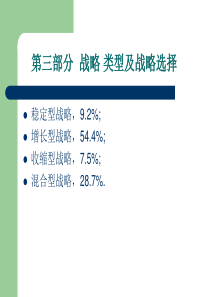
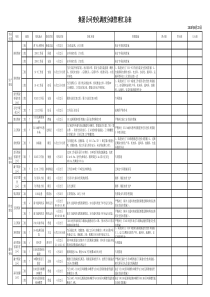
膜泡运输生物工程学院安雯蛋白质分选类型跨膜转运膜泡运输选择性门控转运细胞质基质中的蛋白质转运膜泡运输膜泡运输是蛋白运输的一种特有方式,普遍存在于真核细胞中。蛋白质通过不同类型的转运小泡从糙面内质网合成部位转运至高尔基体进而分选至细胞的不同部位。转运过程不仅涉及蛋白本身的修饰、加工和装配,还涉及到多种不同运输小泡的定向转运及膜泡出芽与融合等复杂的调控过程。三种主要的运输小泡①网格蛋白有被小泡高尔基体TGN质膜、包内体或溶酶体和植物液泡②COPⅡ有被小泡内质网高尔基体③COPⅠ有被小泡回收、转运内质网逃逸蛋白返回内质网网格蛋白有被小泡结构网格蛋白由相对分子质量为180kDa的重链和相对分子质量为35~40kDa的轻链组成二聚体。二聚体三联体骨架六(或五)边形网格蛋白亚基网格蛋白小泡网格蛋白的结构形成过程:●1、被膜小窝的形成网格蛋白被膜小窝是网格蛋白小泡形成过程中的一个中间体。在胞吞过程中,吞入物(配体)先同膜表面特异受体结合,然后网格蛋白装配的亚基结合上去,使膜凹陷成小窝状。由于这种小窝膜外侧结合有许多网格蛋白,故称为网格蛋白被膜小窝。●网格蛋白小泡的形成在形成了网格蛋白被膜小窝之后,很快通过出芽的方式形成小泡,即网格蛋白小泡,小泡须在发动蛋白的作用下与质膜割离。由于此时的小泡外面有网格蛋白包被,故称为被膜小泡。●无被小泡的形成网格蛋白小泡形成之后,很快脱去网格蛋白的外被,成为无被小泡。在真核细胞中有一种分子伴侣Hsc70催化披网格蛋白小泡的外被去聚合形成三腿复合物,并重新用于披网格蛋白小泡的装配。网格蛋白小泡的形成过程在高尔基体TGN区网格蛋白有被小泡形成的示意图COPⅡ有被小泡具有对转运物质的选择性并使之浓缩。原因:①能识别并结合跨膜内质网蛋白胞质面一端的信号序列;②跨膜内质网蛋白的腔面一端作为受体与ER腔中的可溶性蛋白(如分泌蛋白)结合。Sar蛋白是一种小的GTP结合蛋白,作为分子开关起调节作用,主要是调节膜泡包被的装配与去装配。当GDP被GTP取代,则激活Sar蛋白并导致它与内质网结合,同时引发其它包被蛋白组分在ER膜上装配、出芽,随即形成COPⅡ有被小泡。COPI有被小泡内质网向高尔基体输送运输小泡时,一部分自身的蛋白质也不可避免的被运送到了高尔基体,如不进行回收则内质网因为磷脂和某些蛋白质的匮乏而停止工作。内质网通过两种机制维持蛋白质的平衡:一是转运泡将应被保留的驻留蛋白排斥在外,例如有些驻留蛋白参与形成大的复合物,因而不能被包装在出芽形成的转运泡中,结果被保留下来;二是通过对逃逸蛋白的回收机制,使之返回它们正常驻留的部位。内质网的正常驻留蛋白,它们在C端含有一段回收信号序列,如果它们被意外地逃逸进入转运泡从内质网运至高尔基体cis面,则cis面的膜结合受体蛋白将识别并结合逃逸蛋白的回收信号,形成COPI衣被小泡将它们返回内质网。内质网腔中的蛋白,如蛋白二硫键异构酶和协助折叠的分子伴侣,均具有典型的回收信号Lys-Asp-Glu-Leu(KDEL)。内质网的膜蛋白(如SRP受体)在C端有一个不同的回收信号,通常是Lys-Lys-X-X(KKXX,X:任意氨基酸),同样可保证它们的回收。KDEL序列引自MolecularBiologyoftheCell.4thed.2002膜泡运输的定向机制选择性融合是保证细胞内定向膜流的因素之一。各类运输小泡之所以能够被准确地和靶膜融合,是因为运输小泡表面的标志蛋白能被靶膜上的受体识别,其中涉及识别过程的两类关键性的蛋白质是SNAREs(solubleNSFattachmentproteinreceptor)和Rabs(targetingGTPase)。其中SNARE介导运输小泡特异性停泊和融合,Rab的作用是使运输小泡靠近靶膜。SNAREsSNAREs的作用是保证识别的特异性和介导运输小泡与目标膜的融合,位于运输小泡上的叫作v-SNAREs,位于靶膜上的叫作t-SNAREs。v-SNAREs和t-SNAREs都具有一个螺旋结构域,能相互缠绕形成跨SNAREs复合体(trans-SNAREscomplexes),并通过这个结构将运输小泡的膜与靶膜拉在一起,实现运输小泡特异性停泊和融合。除了SNARE之外,还有其他的蛋白参与运输泡与目的膜的融合。t-和v-SNARE
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码








 小牛试刀
小牛试刀
本文标题:膜泡运输
链接地址:https://www.777doc.com/doc-237138 .html