您好,欢迎访问三七文档
当前位置:首页 > 行业资料 > 交通运输 > 哺乳动物细胞表达系统
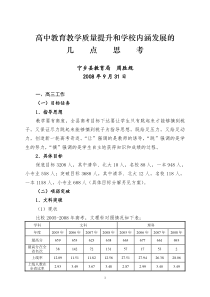
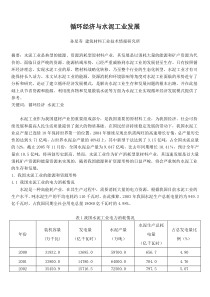
哺乳动物细胞表达系统按照宿主细胞的类型,可将基因表达系统大致分为原核、酵母、植物、昆虫和哺乳动物细胞表达系统。与其它系统相比,哺乳动物细胞表达系统的优势在于能够指导蛋白质的正确折叠,提供复杂的N型糖基化和准确的O型糖基化等多种翻译后加工功能,因而表达产物在分子结构、理化特性和生物学功能方面最接近于天然的高等生物蛋白质分子。从最开始以裸露DNA直接转染哺乳动物细胞至今的30余年间,哺乳动物细胞表达系统不仅已成为多种基因工程药物的生产平台,在新基因的发现、蛋白质的结构和功能研究中亦起了极为重要的作用。本文主要从表达系统及其两个组成部分——表达载体和宿主细胞等方面,简要介绍哺乳动物细胞表达系统和相关的研究进展。研究现状①部分蛋白在哺乳动物细胞中的表达已从实验室研究迈向生产或中试生产阶段。②已有许多重要的蛋白及糖蛋白利用哺乳动物细胞系统表达和大量制备、生产。如人组织型血纤蛋白酶原激活因子、凝血因子Ⅷ、干扰素、乙肝表面抗原、红血球生成激素、人生长激素、人抗凝血素Ⅲ,集落刺激因子等。有些产品已投入临床应用或试用。③虽然经过多年努力,哺乳动物细胞表达系统的表达水平有大幅度增高,但从整个水平上看仍偏低,一般处在杂交瘤细胞单克隆抗体蛋白产率的下限,即1-30μg/l08细胞/24小时。有人认为其限速步骤可嚣是在工程细胞中(对于重组蛋白来讲,常是异源的),重组蛋白的分泌效率较低。1表达载体1.1表达栽体的类型哺乳动物细胞表达外源重组蛋白可利用质粒转染和病毒载体的感染。利用质粒转染获得稳定的转染细胞需几周甚至几个月时间,而利用病毒表达系统则可快速感染细胞,在几天内使外源基因整合到病毒载体中,尤其适用于从大量表达产物中检测出目的蛋白。根据进入宿主细胞的方式,可将表达载体分为病毒载体与质粒载体。病毒载体是以病毒颗粒的方式,通过病毒包膜蛋白与宿主细胞膜的相互作用使外源基因进入到细胞内。常用的病毒载体有腺病毒、腺相关病毒、逆转录病毒、semliki森林病毒(sFv)载体等。另外,杆状病毒载体应用于哺乳动物细胞的表达在近几年颇受重视,这是因为它与其它病毒载体相比有特有优势,如可通过昆虫细胞大量制备病毒颗粒;可感染多种哺乳动物细胞,但在细胞内无复制能力,生物安全度高;可插入高达38kb的外源基因等。质粒载体则是借助于物理或化学的作用导人细胞内。依据质粒在宿主细胞内是否具有自我复制能力,可将质粒载体分为整合型和附加体型载体两类。整合型载体无复制能力,需整合于宿主细胞染色体内方能稳定存在,如SV40病毒载体、反转录病毒载体和游离型如痘苗病毒、腺病毒载体。利用Sindbisvirus(SV)、ScmlikiForestvirus(sFV)和痘苗病毒载体感染哺乳动物细胞表达的蛋白在结构与功能上与天然哺乳动物来源的蛋白更相似。Liljestrom等利用SFV病毒载体感染哺乳动物细胞获得的外源蛋白占细胞总蛋白的;而附加体型载体则是在细胞内以染色体外可自我复制的附加体形式存在。整合型载体一般是随机整合入染色体,其外源基因的表达受插入位点的影响,同时还可能会改变宿主细胞的生长特性。相比之下,附加体型载体不存在这方面的问题,但载体DNA在复制中容易发生突变或重排。附加体型载体在胞内的复制需要两种病毒成分:病毒DNA的复制起始点(ori)及复制相关蛋白。根据病毒成分的来源不同,附加体型表达载体主要分为4大类,表2对这几类附加体载体进行了简要的概括。载体的选择取决于外源基因的导人方式和其调控元件是否有利于转录和翻译。真核生物基因高表达载体必须具有如下调控元件:①原核DNA序列,包括能在大肠杆菌中自身复制的复制子,便于筛选含萤组细菌的抗生素抗性基躅,以及便于目的基因插入的限制性酶切位点。目前采州的哺乳动物细胞表达戟体大都带有来自pBR322的衍生质粒如pX[3、pBRd和pM[的原核序列;②启动子和增强子;③剪接信号;④终止信号和poIyA加尾信号。为了将含目的基因的载体导入哺乳动物功物细胞.还必须加入遗传选择标记。常用的标记基因有胸腺激酶(tk)基因、二氢叶酸还原酶(dh]r)基因、新霉素(neo)抗性基因、氯霉素乙酰基转移酶(cat)基因等dhfr还可作为共扩增基因使外源基因的表达产物增加。当培养基中逐新增加氨甲蝶呤(MTX)的浓度时,随着细胞对MTX抗体的增加。dhfr基因与外源基因均明显扩增。据文献报道,在不断提高的选择压力下,dhfr及侧翼序列能扩增至上千个拷贝,大大增加目的基因的表达水平。1.2表达载体的结构元件哺乳动物细胞表达载体的必要元件包括:一个高活性的启动子、转录终止序列和一个有效的mRNA翻译信号。可视实验需要加入标志基因、复制起始点序列、内部核糖体进入位点等。基于启动子/增强子是表达载体中最重要的元件,我们这里仅对它做简要介绍。外源基因在哺乳动物细胞中的表达与多种因素有关,主要是启动子和增强子的强弱以及它们之间的搭配。启动子需包含两个识别序列:mRNA转录起始点和TATA盒。TATA盒位于转录起始值点上游25--30bp处,是引导RNA聚合酶在正确起始位点转录所必需的序列,即保证转录的精确起始。其他上游启动子元件常位于TATA盒上游100~200bp之间。其功能是调节转录的起始频率和提高转录效率。启动于和增强子受细胞类型的限制,在不同的细胞系中有很大不同,因此需根据宿主细胞的娄型选择不同的启动子和增强子以便于目的基因的高效表达。目前常用的强启动子包括人巨细胞病毒早期启动子(CMV-IE)、人延伸因子1-亚基启动子和Rous肉瘤长末端重复序列;Invitrogen公司开发的pcDNA、pEF和pRL三种系列载体即分别是以这三种启动子驱动目的基因的表达。近年来又发现了一些新的强启动子:如人leukosialin基因同和鼠3-磷酸甘油激酶l(PGKI)基因启动子,活性与CMV-IE相当;人编在蛋白(ubiquitin)C基因启动子不仅具有较高的活性,而且比CMV-IE、PGK1等启动子有更广泛的宿主细胞范围,几乎在转基因小鼠的所有组织细胞中都具有较高活性。核内小RNA(snRNA)启动子亦具有与CMV-IE相近的活性,此前认为该启动子的转录产物不具有正常mRNA的修饰过程和功能,虽然转录同样是由RNA聚合酶Ⅱ完成;但Bartlett等的研究显示,UlsnRNA启动子合成的lacZ基因RNA可被正确地在5’端加帽和3’端加polyA尾,并与核糖体结合引导Iacz蛋白的翻译。常用的增强子有Rous肉癯病毒基因长末端重复序列和人巨细胞病毒增强子。james等利用糖皮质类周醇诱导的鼠乳房瘤病毒(MMTV)启动子和鼠乳房痛病毒长末端重复序列(MMTV-LTR),在CHO细胞中表达分泌的碱性磷酸酶,产量为利用常规的SV40和CMV启动子的10倍,超过0.4mg/ml。构建杂合的启动子是获得新启动子的一个重要途径,比如由人ubiquitinC启动区序列与CMV增强子组成的杂合启动子、由SV40早期启动子和人I型T淋巴细胞病毒LTR中的增强子序列(R-U5片段)组成的SR-α启动子的活性均与CMV-IE相当;而由鸡β-肌动蛋白启动子和CMV增强子序列构成的杂合启动子不仅活性比CMV—IE高,而且具有更为广谱的宿主细胞范围。Novagen公司的pBacMam表达系统即是以该杂合启动子驱动目的基因的转录。另一类杂合启动子是由活性很低的启动子与数个转录激活因子结合位点串联而成,这些位点与转录激活因子的结合受细胞外小分子药物的调控;这类启动子主要用作基因的诱导表达。如Invitrogen公司的ecdysone诱导表达系统,其负责目的基因转录的启动子是由热休克蛋白启动子核心序列(minimalpromotor)和5个E/GRE元件组成;Clontech公司开发的Tet-off系统中的启动子则由CMV启动子的核心序列和7个Tet阻遏蛋白结合位点组成。这些启动子在诱导前后活性可相差4个数量级。另外,在哺乳动物细胞中已发现存在大量在低氧环境中可诱导转录的基因,如编码红细胞生成素(EPO)转铁蛋白、血红素加氧酶-1等的基因,它们都有一个共同的顺式作用元件(CGTG),有利于在5’或3’侧翼区的低氧诱导低氧诱导作用因子-1(HIF-1)和低氧反应增强子(HRE)结合,激活靶基因的转录,在低氧浓度下可使重组蛋白大量表达。据报道红细胞生成素启动子在1%氧浓度比在21%氧浓度下活陛增强100倍。这又是一种新的提高表达量的作用机制。2宿主细胞哺乳动物细胞表达外源蛋白最初是将抗体基因重新导人淋巴细胞中由病毒(如SV40)或lgG的启动子增强子引导。产生的抗体具有相应的结合能力和数应功能,但表达量很低。常用的非淋巴细胞类有中国仓鼠卵巢(CHO)细胞、小仓鼠肾(BHK)细胞、COS细胞、小鼠NSO胸腺瘤细胞和小鼠骨髓瘤SP2/0细胞等。不同宿主细胞表达的重组蛋白其稳定性和蛋白糖基化类型不同,需根据要表达的目的蛋白选择最佳的宿主细胞。COS细胞是进行外源基因瞬时表达时用途最广的宿主,其重组载件易于组建,便于使用,而且对插入DNA的量或者采用基因组DNA序列的情况都没有什么限制,便于通过检测表达情况来确证cDNA的阳性克隆,也利于快速分析引入克隆化cDNA序列中的突变。CHO细胞则利于外源基目的稳定整合,易于大规模培养,能在无血清和蛋白的条件下生比,是用于真核生物基因表达软为成功的宿主细胞。已用于多种复杂的重组蛋白的生产,但其产量较低,一般仅占细胞蛋白的2.5%,而用细菌表达可获得占总蛋白50%的蛋白表达水平,但大肠杆菌表达的动物蛋白能进行正确的翻译后加工如糖基化和三维结构的形成,不具有与天然抗体相似的功能活性,且在人体内易于清除。如果需要表达具有生物学功能的膜蛋白或分泌型蛋白,例如细胞表面的受体或细胞外的激素和酶,则不能在原棱细胞中表达。而哺乳动物细胞表达的蛋白则具有天然蛋白的生物学活性。为提高哺乳动物细胞的蛋白表达量,需选择合适的表达载体和有效的启动子和增强子。近几年也陆续发现了几种新的有较大应用价值的细胞株:如来源于MadIin-Darby犬肾的高分化内皮细胞株(MDCK),Pei等用该细胞株表达分泌型的基质金属蛋白酶MMPI3,发现高表达的阳性细胞克隆可占转染细胞的5%~l0%,其中一个克隆的表达量可占细胞上清总蛋白的l5%~20%,在细胞单层贴壁培养情况下表达量达10ms/L。作者认为如此高的表达量可能与MDCK细胞的特性有关,因为同样的载体在CH0细胞中仅得到低水平的表达。在球蛋白基因簇上游50kb处发现有一区域在红细胞系的细胞中起调控基因表达的作用,称为位点控制区域(LCR),LCR可调整附近区域染色体的结构并增强球蛋白的转录活性。一些外源基因如thy-l和TK基因与LCR相连后均可在红系细胞中高表达,而且表达水平与载体的插入位点无关,这一特性使得红系细胞颇受关注。MEL—E9是目前常用于重组蛋白表达的红系细胞株,它来源于感染了SSF病毒(spleenfocusformingvirus)的N小鼠的脾细胞。对现有细胞株选择性地进行遗传改造是提高重组蛋白表达水平的重要手段。BHK/vl6细胞株是稳定表达单纯疱疹病毒(HSV)VPI6蛋白的BHK细胞,由于VPI6的转录激活作用,载体中的HSV早期启动子在该工程细胞中有很高的活性;已用该细胞株获得了包括人组织纤溶酶原激活剂、生长因子等在内的多种蛋白的高效稳定表达。基于腺病毒EIA蛋白可反式激活CMV启动子,Cockett等建立了稳定表达EIA的CHO细胞株,在该细胞中重组前胶原酶的产量与CHO细胞相比增加了l2倍;该细胞在多种抗体的表达中亦取得满意的结果。对细胞其它特性的遗传改造,包括细胞生长周期的调控、细胞的抗凋亡能力、细胞贴壁能力、蛋白糖基化的模式及细胞乳酸的合成等方面,亦有相关报道,其实际应用价值有待得到更多实验数据的支持。3表达系统根据目的蛋白表达的时空差异,可将表达系统分为瞬时、稳定和诱导表达系统。瞬时表达系统是指宿主细胞在导入表达载体后不经选择培养,载体DNA随细胞分裂而逐渐丢失,目的蛋白的表达时限短暂;瞬时表达系统的优点是简捷,实
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码








 gsheep
gsheep
本文标题:哺乳动物细胞表达系统
链接地址:https://www.777doc.com/doc-3407851 .html