您好,欢迎访问三七文档
当前位置:首页 > 商业/管理/HR > 经营企划 > 核酸外切酶、内切酶、聚合酶、连接酶
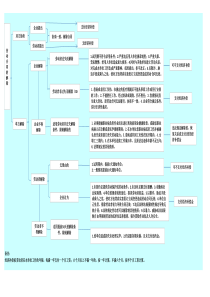
核酸外切酶(exonuclease)核酸外切酶是一类能从多核苷酸链的一端开始按序催化水解3、5-磷酸二酯键,降解核苷酸的酶。其水解的最终产物是单个的核苷酸(DNA为dNTP,RNA为NTP)。按作用的特性差异可以将其分为单链的核酸外切酶和双链的核酸外切酶。单链的核酸外切酶包括大肠杆菌核酸外切酶Ⅰ(exoⅠ)和核酸外切酶Ⅶ(exoⅦ)。核酸外切酶Ⅶ(exoⅦ)能够从5′-末端或3′-末端呈单链状态的DNA分子上降解DNA,产生出寡核苷酸短片段,而且是唯一不需要Mg2+离子的活性酶,是一种耐受性很强的核酸酶。核酸外切酶Ⅶ(exoⅦ)可以用来测定基因组DNA中一些特殊的间隔序列和编码序列的位置。它只切割末端有单链突出的DNA分子。双链的核酸外切酶包括大肠杆菌核酸外切酶Ⅲ(exoⅢ)、λ噬菌体核酸外切酶(λexo)以及T7噬菌体基因6核酸外切酶等。大肠杆菌核酸外切酶Ⅲ(exoⅢ)具有多种催化功能,可以降解双链DNA分子中的许多类型的磷酸二酯键。其中主要的催化活性是催化双链DNA按3′→5′的方向从3′-OH末端释放5′-单核苷酸。大肠杆菌核酸外切酶Ⅲ(exoⅢ)通过其3′→5′外切酶活性使双链DNA分子产生出单链区,经过这种修饰的DNA再配合使用Klenow酶,同时加进带放射性同位素的核苷酸,便可以制备特异性的放射性探针。λ噬菌体核酸外切酶(λexo)最初是从感染了λ噬菌体的大肠杆菌细胞中纯化出来的。这种酶催化双链DNA分子从5′-P末端进行逐步的水解释放出5′-单核苷酸。但不能降解5′-OH末端。内切酶(incisionenzyme)这是一类能从DNA分子中间水解磷酸二酯键,从而切断双链DNA的核酸水解酶。它们不同于一般的脱氧核糖核酸酶(DNase),它们的切点大多很严格,要求专一的核苷酸顺序识别顺序内切酶主要分成三大类。第一类内切酶能识别专一的核苷酸顺序,并在识别点附近的一些核苷酸上切割DNA分子中的双链,但是切割的核苷酸顺序没有专一性,是随机的。这类限制性内切酶在DNA重组技术或基因工程中没有多大用处,无法用于分析DNA结构或克隆基因。这类酶如EcoB、EcoK等。第二类内切酶能识别专一的核苷酸顺序,并在该顺序内的固定位置上切割双链。由于这类限制性内切酶的识别和切割的核苷酸都是专一的。所以总能得到同样核苷酸顺序的DNA片段,并能构建来自不同基因组的DNA片段,形成杂合DNA分子。因此,这种限制性内切酶是DNA重组技术中最常用的工具酶之一。这种酶识别的专一核苷酸顺序最常见的是4个或6个核苷酸,少数也有识别5个核苷酸以及7个、9个、10个和11个核苷酸的。如果识别位置在DNA分子中分布是随机的,则识别4个核苷酸的限制性内切酶每隔46(4096)个核苷酸就有一个切点。人的单倍体基因组据估计为3×199核苷酸,识别4个核苷酸的限制性内切酶的切点将有(3×109/2.5×102)约107个切点,也就是可被这种酶切成107片段,识别6个核苷酸的限制性内切酶也将有(3×109/4×103)约106个切点。第三类内切酶的识别顺序是一个回文对称顺序,即有一个中心对称轴,从这个轴朝二个方向“读”都完全相同。这种酶的切割可以有两种方式。一是交错切割,结果形成两条单链末端,这种末端的核苷酸顺序是互补的,可形成氢键,所以称为粘性末端。粘性末端的意义:(1)连接便利:不同的DNA双链,只要粘性末端碱基互补就可以连接,这比连接两个平齐末端容易的多;同一个DNA分子内连接,通过两个相同的粘性末端可以连接成环状分子。(2)5’末端标记:凸出的5’末端可以用DNA多核苷酸激酶进行32P标记。凸出的3’端可以通过末端转移酶添加几个多聚核苷酸的尾巴(如AAA或TTT等)造成人工粘性末端(3)补平成平齐末端粘性末端可以用DNA聚合酶补平成平齐末端如EcoRI的识别顺序为:↓|5’……GAA|TTC……3’3’……CTT|AAG……5’|↑垂直虚线表示中心对称轴,从两侧“读”核苷酸顺序都是GAATTC或CTTAAG,这就是回文顺序(palindrome)。实线剪头表示在双链上交错切割的位置,切割后生成5’……G和AATTC……3’、3’……CTTAA和G……5’二个DNA片段,各有一个单链末端,二条单链是互补的,可通过形成氢键而“粘合”。另一种是在同一位置上切割双链,产生平头末端。例如HaeⅢ的识别位置是:↓5’……GG↓CC……3’3’……CC↓GG……’↑在箭头所指处切割,产生的两个DNA片段是:5’……GGCC……3’和3’……CCGG……5’有时候两种限制性内切酶的识别核苷酸顺序和切割位置都相同,差别只在于当识别顺序中有甲基化的核苷酸时,一种限制性内切酶可以切割,另一种则不能。例如HpaⅡ和MspⅠ的识别顺序都是5’……GCGG……3’,如果其中有5’-甲基胞嘧啶,则只有HpaⅡ能够切割。这些有相同切点的酶称为同切酶或异源同工酶(isoschizomer)。第三类内切酶也有专一的识别顺序,但不是对称的回文顺序。它在识别顺序旁边几个核苷酸对的固定位置上切割双链。但这几个核苷酸对则是任意的。因此,这种限制性内切酶切割后产生的一定长度DNA片段,具有各种单链末端。这对于克隆基因或克隆DNA片段没有多大用处。DNA聚合酶(DNApolymerase)DNA聚合酶(DNApolymerase)是细胞复制DNA的重要作用酶。DNA聚合酶,以DNA为复制模板,从将DNA由5'端点开始复制到3'端的酶。DNA聚合酶的主要活性是催化DNA的合成(在具备模板、引物、dNTP等的情况下)及其相辅的活性。DNA聚合酶的共同特点是:(1)需要提供合成模板;(2)不能起始新的DNA链,必须要有引物提供3'-OH;(3)合成的方向都是5'→3'(4)除聚合DNA外还有其它功能。功能[1]聚合作用:在引物RNA'-OH末端,以dNTP为底物,按模板DNA上的指令由DNApolⅠ逐个将核苷酸加上去,就是DNApolⅠ的聚合作用。酶的专一性主要表现为新进入的脱氧核苷酸必须与模板DNA配对时才有催化作用。dNTP进入结合位点后,可能使酶的构象发生变化,促进3'-OH与5'-PO4结合生成磷酸二酯键。若是错误的核苷酸进入结合位点,则不能与模板配对,无法改变酶的构象而被3'-5'外切酶活性位点所识别并切除之。[2]3'→5'外切酶活性──校对作用:这种酶活性的主要功能是从3'→5'方向识别和切除不配对的DNA生长链末端的核苷酸。当反应体系中没有反应底物dNTP时,由于没有聚合作用而出现暂时的游离现象,从而被3'→5'外切酶活性所降解。如果提高反应体系的温度可以促进这种作用,这表明温度升高使DNA生长链3'末端与模板发生分离的机会更多,因而降解作用加强。当向反应体系加入dNTP,而且只加放与模板互补的上述核苷酸才会使这种外切酶活性受到抑制,并继续进行DNA的合成。由此推论,3'→5'外切酶活性的主要功能是校对作用,当加入的核苷酸与模板不互补而游离时则被3'→5'外切酶切除,以便重新在这个位置上聚合对应的核苷酸。在某些T4噬菌体突变株中DNA复制的真实性降低,而易发生突变,从此突变株分离得到的T4DNA聚合酶的3'→5'外切酶活性很低。相反,另外一些具有抗突变能力的T4突变株中的T4DNA聚合酶的3'→5'外切酶活性比野生型高得多,因此,其DNA复制真实性好,变异率低。可见,3'→5'外切酶活性对DNA复制真实性的维持是十分重要的。[3]5'→3'外切酶活性──切除修复作用:5'→3'外切酶活性就是从5'→3'方向水解DNA生长链前方的DNA链,主要产生5'-脱氧核苷酸。这种酶活性只对DNA上配对部分(双链)磷酸二酯键有切割活力作用,方向是5'→3'。每次能切除10个核苷酸,而且DNA的聚合作用能刺激5'→3'外切酶活力达10倍以上。因此,这种酶活性在DNA损伤的修复中可能起着重要作用。对冈崎片段5'末端RNA引物的去除依赖此种外切酶活性。[4]焦磷酸解作用:DNApolⅠ的这种活性可以催化3'末端焦磷酸解DNA分子。这种作用就是无机焦磷酸分解DNA生长链,可以认为是DNA聚合作用的逆反应,而且这种水解DNA链作用需要有模板DNA的存在。(dNMP)n+XPPi←(dNMP)n-x+X(dNPPP)→DNA[5]焦磷酸交换作用:催化dNTP末端的PPi同无机焦磷酸的交换反应。反应式为32P32Pi+dNPPP←dNP32P32P+PPi→DNA最后两种作用,都要求有较高浓度的PPi,因此,在体内由于没有足够高的PPi而无重要意义。DNApolⅠ的DNA聚合酶活性和5'→3'外切酶活性协同作用,可以使DNA链上的切口向前推进,即没有新的DNA合成,只有核苷酸的交换。这种反应叫缺口平移(Nicktranslation)。当双链DNA上某个磷酸二酯键断裂产生切口时,DNApoIⅠ能从切口开始合成新的NDA链,同时切除原来的旧链。这样,从切口开始合成了一条与被取代的旧链完全相同的新链。如果新掺入的脱氧核苷酸三磷酸为α-32P-dNTP,则重新合成的新链即为带有同位素标记的DNA分子,可以用作探针进行分子杂交实验。大肠杆菌DNA聚合酶1、大肠杆菌DNA聚合酶Ⅰ(DNApolymeraseⅠ,DNApolⅠ)这是1956年由ArthurKornberg首先发现的DNA聚合酶,又称Kornber酶。此酶研究得清楚而且代表了其他DNA聚合酶的基本特点。(1)理化性质:纯化的DNApolⅠ由一条多肽链组成,约含1000个氨基酸残基,MW为109KD。分子含有一个二硫键和一个-SH基。通过二个酶分子上的-SH基与Hg2+结合产生二聚体,仍有活性。每个酶分子中含有一个Zn2+,在DNA聚合反应起着很重要的作用。每个大肠杆菌细胞中含有约400个分子,每个分子每分钟在37℃下能催化667个核苷酸掺入正在生长的DNA链。经过枯草杆菌蛋白酶处理后,酶分子分裂成两个片段,大片段分子量为76KD,通常称为Klenow片段,小片段的分子量为34KD。此酶的模板专一性和底物专一性均较差,它可以用人工合成的RNA作为模板,也可以用核苷酸为底物。在无模板和引物时还可以从头合成同聚物或异聚物。DNAPolⅠ在空间结构上近似球体,直径约65A。在酶的纵轴上有一个约20A的深沟(cleft),带有正电荷,这是该酶的活性中心位置,在此位置上至少有6个结合位点:[1]模板DNA结合位点;[2]DNA生长链或引物结合位点;[3]引物末端结合位点,用以专一引物或DNA生长链的3'-OH;[4]脱氧核苷三磷酸结合位点;[5]5'→3'外切酶活性位点,用以结合生长链前方的5'-端脱氧核苷酸并切除之;[6]3'→5'外切酶活性位点,用以结合和切除生长链上未配对的3'-端核苷酸。大肠杆菌DNA聚合酶I大片段(Klenow)Klenow酶的基本用途:(1)补平由核酸内切酶产生的5’粘性末端(2)DNA片段的同位素末端标记(3)cDNA第二链的合成(4)双脱氧末端终止法测定DNA序列2、大肠杆菌DNA聚合酶Ⅱ(DNApolⅡ):DNA聚合酶Ⅰ缺陷的突变株仍能生存,这表明DNApolⅠ不是DNA复制的主要聚合酶。人们开始寻找另外的DNA聚合酶,并于1970年发现了DNApolⅡ。此酶MW为120KD,每个细胞内约有100个酶分子,但活性只有DNApolⅠ的5%。该酶的催化特性如下:(1)聚合作用:该酶催化DNA的聚合,但是对模板有特殊的要求。该酶的最适模板是双链DNA而中间有空隙(gap)的单链DNA部分,而且该单链空隙部分不长于100个核苷酸。对于较长的单链DNA模板区该酶的聚合活性很低。但是用单链结合蛋白(SSBP)可以提高其聚合速率,可达原来的50-100倍。(2)该酶也具有3'→5'外切酶活性,但无5'→3'外切酶活性。(3)该酶对作用底物的选择性较强,一般只能将2-脱氧核苷酸掺入到DNA链中。(4)该酶不是复制的主要聚合酶,因为此酶缺陷的大肠杆菌突变株的DNA复制都正常。可能在DNA的损伤修复中该酶起到一定的作用。3、大肠杆菌DNA聚合酶Ⅲ(DNAp
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码
![[能源行业]建设项目基本情况-年产50万吨沥青混凝土生产线建设项目(DOC 45页)](/doc-257037.png)








 olo0001
olo0001
本文标题:核酸外切酶、内切酶、聚合酶、连接酶
链接地址:https://www.777doc.com/doc-3471887 .html