当前位置:首页 > 商业/管理/HR > 信息化管理 > 白介素IL-6信号转导及其通路研究概述
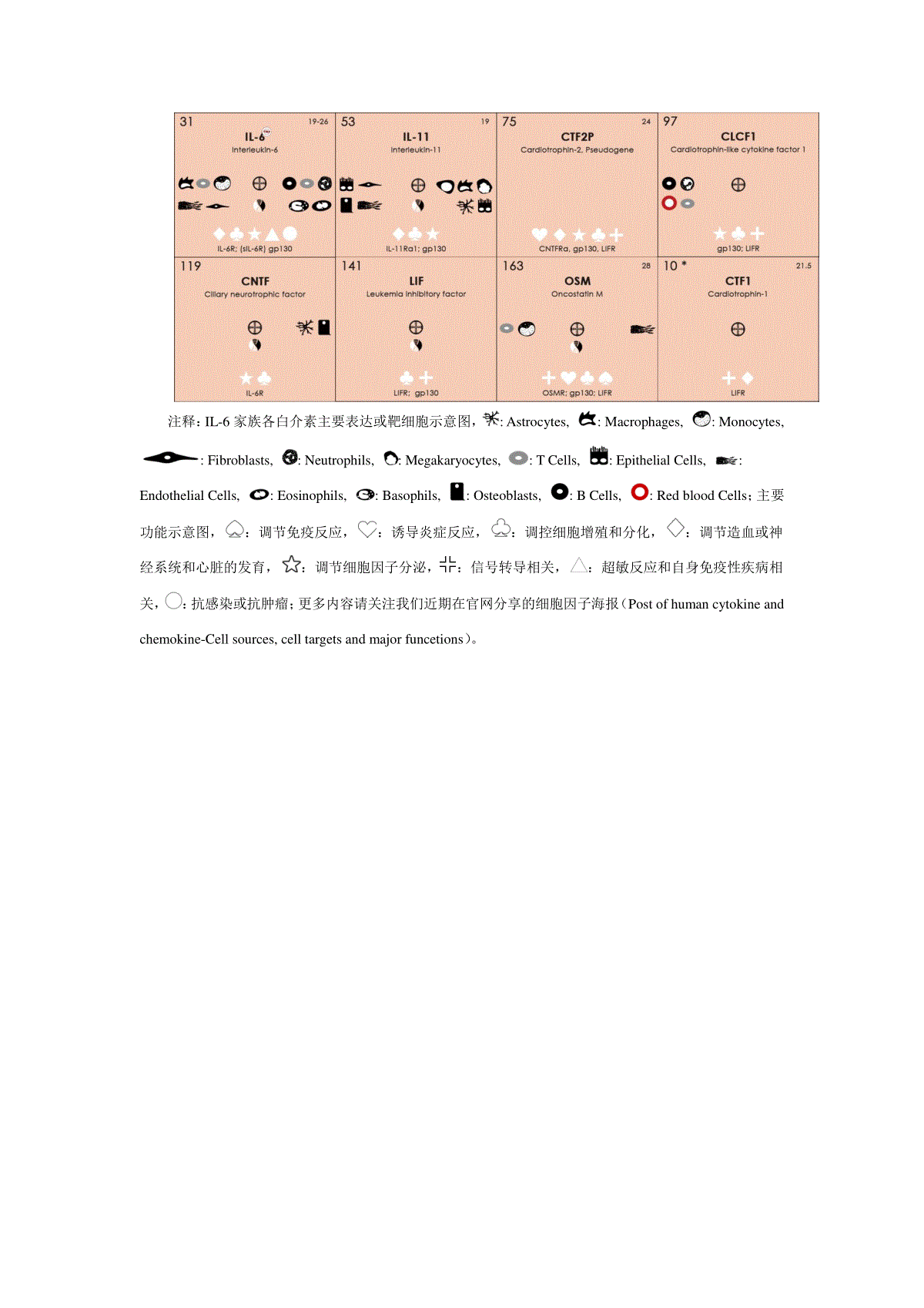
白介素IL-6信号转导及其通路研究概述细胞因子是一类参与免疫系统的细胞之间通信的蛋白质,除此之外,许多细胞因子在免疫系统之外也具有调节功能。1986年白介素IL-6作为B细胞刺激因子被Kishimoto组分子克隆。IL-6在免疫系统外的活性还有肝细胞刺激因子和骨髓细胞分化诱导蛋白。白介素IL-6含有184个氨基酸,属于糖基化蛋白质。IL-6可以由多种类型细胞合成和分泌,包括单核细胞、T细胞、成纤维细胞和内皮细胞。IL-6结合受体有两种,一种是特异性受体IL-6R(80kDaI型跨膜蛋白),另一种是gp130,是IL-6家族细胞因子的所有成员的常见受体亚单位。gp130可以在所有细胞表达,但IL-6R的表达受到更多的限制,主要发现于肝细胞、嗜中性粒细胞、单核细胞和CD4+T细胞。白介素IL-6受体gp130的二聚化会导致两种细胞内信号通路的启动:经典信号通路和反式信号通路(见下文)。白介素IL-6的受体IL-6R可以在细胞膜经过蛋白质水解,形成可溶性的IL-6R(sIL-6R),在人类中,也可以在翻译阶段进行剪接mRNA,进而产生sIL-6R。在经典信号通路中,IL-6与膜上的IL-6R结合,随后与结合在细胞膜上的gp130结合,启动细胞内信号传导。在IL-6反式信号通路中,IL-6与sIL-6R结合,IL-6和sIL-6R的复合物与细胞膜结合的gp130结合,从而引发细胞内信号。白介素IL-6是最重要的炎症细胞因子之一。IL-6在通过膜结合和可溶性受体的信号传导中是独特的。有趣的是,这两种途径的生物学后果有很大差异,通过膜结合受体的经典IL-6信号通路主要是再生和保护性的,可溶性IL-6R的IL-6反式信号通路是促炎症的。响应于受体激活的IL-6的细胞内信号传导是通过STAT依赖和STAT独立的信号模块,其由复杂的调节网络调节。IL-6的复杂生物学对该细胞因子的治疗靶向具有影响。白介素IL-6胞内信号通路可以简单的概述为:IL-6与受体复合物结合后,激活JAK1。JAK1磷酸化gp130细胞质部分内的酪氨酸残基,这些磷酸酪氨酸基序是STAT转录因子,SOCS3反馈抑制剂和衔接蛋白和磷酸酶SHP2的募集位点。SHP2连接到MAPK级联,使Gab1磷酸化,磷酸化的Gab1转移到质膜上,协调正在进行的MAPK和PI3K活化。Src家族激酶独立于受体磷酸化并激活YAP。白介素IL-6信号转导第一步:激活JAK。大多数细胞因子受体缺乏胞内激酶活性,生长因子的受体例外。白介素IL-6胞内信号转导首先激活Janus激酶(JAK),开启酶促反应。通过JAKN末端的同源结构域内(JH)JH7-JH5的FERM-结构域介导,与gp130相互作用。在受体位点,分别含有富含脯氨酸基序和疏水基序的膜近端box1和box2区域,有助于JAK和gp130的相互作用。IL-6与受体的结合后,JAK通过自磷酸化激活,主要在激酶的C-末端JH1-结构域内的激活环的双酪氨酸基序上。附近定位的所谓的假激酶结构域JH2不会发挥酶活性,但对JH1具有调节和抑制作用。白介素IL-6激活Src家族激酶。IL-6信号转导过程中除了激活JAK激酶外,还可以激活Src家族激酶。关于Src激酶的功能首先是在对白血病抑制因子(LIF)信号转导的研究发现的。Src家族激酶Hck与未分化的胚胎干细胞(ES)细胞中的LIF反应,并与gp130相互作用。在多发性骨髓瘤细胞中也证实了Src家族激酶与gp130的相互作用。IL-6诱导Hck和Lyn,与gp130相互作用,Src激酶(HcK,Lyn,Fyn)的随后激活。Hck有助于IL-6信号传导和IL-6依赖性增殖。此外,Hck响应于IL-6介导多位点对接蛋白Gab1的酪氨酸磷酸化。由于Hck主要在造血细胞中表达,因此在非造血细胞中,其他Src激酶是否在IL-6信号传导中占据了Hck的功能,这一点尚待阐明。Src激酶Btk和Tec也参与造血,发现与gp130相互作用。白介素IL-6激活STAT分子。活化的JAK激酶磷酸化gp130细胞质部分内的酪氨酸基序。募集到gp130的最突出的蛋白质是信号转导和转录激活因子(STAT)STAT3和(在一定程度上)STAT1的转录因子。典型的JAK-STAT信号通路,STAT1或STAT3单体与酪氨酸基序结合,STAT因子的酪氨酸磷酸化,酪氨酸磷酸化的STAT1和/或STAT3形成同源二聚体或异源二聚体,随后将STAT二聚体转运入细胞核,在核中,STAT二聚体结合IL-6诱导型基因启动子的增强子元件。白介素IL-6激活独立的STAT3信号通路。除STAT之外,IL-6还激活MAPK级联和PI3K信号通路。STAT3和MAPK激活之间的平衡对于生物体的体内平衡和细胞增殖至关重要。这种平衡由gp130的细胞质酪氨酸基序调节。这种受体基序消除酪氨酸磷酸化,增加IL-6依赖性STAT3活化,降低MAPK激活,导致胃腺瘤、自身免疫性关节炎和增强的Th1细胞因子表达。反之,酪氨酸磷酸化会限制STAT3激活,促进MAPK激活,导致急性期基因表达降低和Th2细胞因子表达增加。最后,两种途径的阻断都会引起急性期反应和血小板减少,而STAT3和MAPK的激活与脾肿大和血小板增多症有关。白介素IL-6信号转导的反向调节。IL-6信号转导通过gp130细胞质部分内的酪氨酸基序759的磷酸化来负调节,可以结合两种蛋白质SHP2和SOCS3,并独立地起作用以抑制JAK-STAT信号传导。而磷酸酶SHP2通过去磷酸化磷酸酪氨酸基序使信号成分失活,JAK抑制剂SOCS3干扰受体活化。SHP2是一种蛋白酪氨酸磷酸酶,在其N-末端区域内含有两个SH2结构域。在非活动状态下,最多的N端SH2结构域覆盖磷酸酶的活性位点。该酶通过SH2结构域与受体或多位点对接蛋白(例如gp130,Gab1)中的磷酸酪氨酸基序结合而激活,随后发挥催化作用。SHP2突变体不能结合gp130或缺乏磷酸酶活性的表达,引起IL-6信号传导增加,证明SHP2对IL-6信号传导的负调控的影响。SOCS3还结合gp130内的磷酸酪氨酸基序759以抑制IL-6信号传导。SOCS3蛋白是JAK激酶的反馈抑制剂,并且被IL-6快速诱导。SOCS3的快速诱导和周转解释了IL-6信号的瞬时动力学。关于持续或短暂IL-6信号的影响的详细的体内研究证明了SOCS3反馈抑制对炎症调节的意义。在缺乏SOCS3反馈抑制作用下,IL-6诱导STAT3的持续激活,并在巨噬细胞中发挥抗炎、IL-10样反应。SOCS3主要通过其结合和抑制JAK激酶催化结构域的短激酶抑制区(KIR)发挥抑制活性。因此,SOCS3、JAK激酶和gp130形成三角形相互作用,其中三种蛋白质中的每一种都与两种其它蛋白质相互作用。IL-6可以通过两种不同的途径起作用,这些途径使用膜结合和可溶性IL-6R。虽然通过膜结合的IL-6R的经典信号传导被认为主要是保护性和再生性的,但IL-6反式信号传导可以被认为代表身体的应激反应以维持体内平衡。由于在自身炎症条件如风湿性关节炎或炎症性肠病中过度照射IL-6活性被认为主要由IL-6反式信号传导介导,因此通过sgp130Fc对该信号通路的特异性阻断可能优于通过中和的全局IL-6阻断针对IL-6或IL-6R的抗体。或者,IL-6信号可以在细胞内被抑制。对STAT独立信号的分子机制的近期见解以及Src家族激酶在IL-6信号转导中的贡献的新知识可能开辟设计干预IL-6信号传导策略的全新视角。然而,开发这些概念需要更深入的了解IL-6信号转导的详细分子机制。附图:白介素IL-6家族因子表达细胞、靶细胞及其主要功能图注释:IL-6家族各白介素主要表达或靶细胞示意图,:Astrocytes,:Macrophages,:Monocytes,:Fibroblasts,:Neutrophils,:Megakaryocytes,:TCells,:EpithelialCells,:EndothelialCells,:Eosinophils,:Basophils,:Osteoblasts,:BCells,:RedbloodCells;主要功能示意图,:调节免疫反应,:诱导炎症反应,:调控细胞增殖和分化,:调节造血或神经系统和心脏的发育,:调节细胞因子分泌,:信号转导相关,:超敏反应和自身免疫性疾病相关,:抗感染或抗肿瘤;更多内容请关注我们近期在官网分享的细胞因子海报(Postofhumancytokineandchemokine-Cellsources,celltargetsandmajorfuncetions)。



 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码










 zrl0127
zrl0127
本文标题:白介素IL-6信号转导及其通路研究概述
链接地址:https://www.777doc.com/doc-4176435 .html