您好,欢迎访问三七文档
当前位置:首页 > 行业资料 > 其它行业文档 > VEGF及其受体的生物学特性及在肿瘤血管生成中的作用-马莉
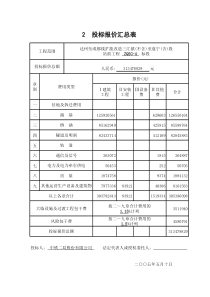
·146·中国优生与遗传杂志2016年第24卷第5期多年来,血管内皮生长因子(vascularendothelialgrowthfactor,VEGF)受到人们的关注,VEGF又称为血管通透因子(VPF),是血管内皮细胞特异性的肝素结合生长因子,它的主要功能是特异性地促进内皮细胞有丝分裂和介导血管形成并增加血管通透性,是一类具有高度生物活性的功能性糖蛋白。VEGF也是许多肿瘤血管生成的核心,主要由多种组织细胞分泌,分布广泛,在肿瘤细胞增殖、凋亡、淋巴道转移等过程中起着至关重要的作用。1 VEGF及其受体结构特点及功能1.1 VEGF的结构及功能VEGF是促内皮细胞分裂原(Mitogen),定位于染色体6p21.3,编码产物是一个分子量为45KD的由8个半胱氨酸残基组成的糖蛋白。VEGF基因家族包括6个成员:VEGF-A(VEGF)、VEGF-B、VEGF-C、VEGF-D、VEGF-E及胎盘生长因子(Placentalgrowthfactor,PlGF),通常所说的VEGF即指VEGF-A,其是一种二硫键结合的二聚体糖蛋白,PIGF之所以命名为胎盘生长因子,就是由于主要高表达在胎盘绒毛膜滋养层中,由于VEGFmRNA不同的剪切方式,产生出分别VEGF121、VEGF145、VEGF165、VEGF189、VEGF206等多种异构体形式,每一个不同的异构体结合相应的受体会因此引起的不同的生物学反应,VEGF是由许多不同类型的组织细胞产生,通过与细胞膜上的受体结合刺激内皮细胞有丝分裂,从而促进新生血管发生、血管生成。此外,VEGF具有高度保守特性,且具有很强的耐热和耐酸能力。VEGF主要有以下几方面的功能:(1)VEGF能促进血管内皮细胞的分裂增殖。(2)VEGF能显著提高血管的通透性,其效应远远强于组胺等血管活性物质。(3)VEGF能促进造血细胞的发育(4)VEGF能促进内皮细胞的浸润和迁移(5)VEGF能促进淋巴内皮细胞生长,并诱导淋巴管的生成。(6)VEGF能改变细胞外基质和促进血管支持物的形成。(7)VEGF能抑制细胞凋亡等[1]。1.2 VEGF受体的结构与功能目前己发现的VEGF受体有4种:VEGFRl(fms-liketyrosinekinase,Flt-1,fms样酪氨酸激酶)、VEGFR2(fetalliverkinase-1,Flk-1/KDR,胎儿肝激酶-1)、VEGFR3(Flt-4)和Neuropilin-1(NRP-1)。其中前三种受体在结构上类似,都属于酪氨酸激酶超家族。VEGFR蛋白由膜外区、跨膜片段及膜内区三部分构成,膜外区由Ig结构域组成,膜内区属酪氨酸激酶区。但是三种受体也有一些区别,比如,VEGFR1(Flt-1)膜外区的第二个Ig结构域是可与配体VEGF结合;VEGFR2(KDR/Flk-1)的第三个Ig结构域专性与配体结合[2];但是VEGFR3(Flt-4)的第五个Ig结构域被二硫键所替代。NRP-1又称神经纤毛蛋白-1,该蛋白是非酪氨酸激酶跨膜糖蛋白的一种,全长由923个氨基酸组成,大小120-130ku,也由胞外区、跨膜区和胞内区三部分组成。VEGF家族对于几种受体有不同的亲和力,VEGFA主要与Flt-1、Flk-1/KDR、NRP-1结合发挥作用,VEGFB选择结合在Flt-1上,PlGF与Flt-1、NRP-1结合,VEGFC和VEGFD可结合在VEGFR2、VEGFR3上。Flt-1与KDR是研究得最多的受体,1.2.1 VEGFR1(Flt-1) VEGFR1(Flt-1)是最早发现的受体,其功能比较复杂。其基因定位于染色体4q12,有研究表明[3],在正常生理情况下受体1和2与VEGF的亲和力不一样,Flt-1与VEGF的亲和力比KDR与VEGF的亲和力高,但酪氨酸酶活性却比KDR低,所以Flt-1调节内皮细胞的活性比KDR低,说明Flt-1对血管的生成可能具有负向调节作用,主要表现为阻止由VEGFR-2介导的基金项目:延安大学校级科研项目(No.YDQ2014-49)VEGF及其受体的生物学特性及在肿瘤血管生成中的作用马 莉(延安大学医学院病原生物学教研室,陕西 延安 716000)摘 要:血管内皮生长因子通过与其受体结合后特异性地促进内皮细胞有丝分裂和介导血管形成并增加血管通透性,也可介导肿瘤血管的形成,肿瘤细胞生长依赖新生血管的形成已经是肿瘤生物学上广为人知的一个方面,肿瘤新生血管在肿瘤的形成、进展、浸润和迁移中起着至关重要的作用。本文就近年来国内外有关VEGF及其受体在肿瘤血管生成方面的作用作一简要综述。关键词:血管内皮生长因子;VEGF受体;肿瘤中图分类号:R730.3 文献标识码:A 文章编号:1006-9534(2016)05-0146-03Theroleofvascularendothelialgrowthfactoranditsreceptorintumorangiogenesis.MALi.(PathogenBiologyTeachingandResearchSection,MedicalSchoolofYan′anUniversity,Yan′an,Shaanxi716000,China)Abstract:Vascularendothelialgrowthfactorpromotesendothelialcellmitosisandangiogenesisandincreasevascularpermeability,whichcanalsomediatetumorangiogenesis,tumorcellgrowthdependsontheformationoftumorbiology.Tumorangiogenesisplaysanimportantroleintumorformation,progression,invasionandmigration.Inthispaper,theeffectsofVEGFanditsreceptorintumorangiogenesiswerereviewed.Keywords:VEGF;VEGFR;TumorDOI:10.13404/j.cnki.cjbhh.2016.05.063·147·中国优生与遗传杂志2016年第24卷第5期信号通路。但是在某些病理情况下,VEGFR1的作用主要是诱导VEGF与VEGFR2结合,从而发挥VEGF促血管形成的生物学作用,起到正向调控VEGF的作用。Hiratsuka等[4]研究认为,缺少VEGFR1膜内区域和跨膜区的小鼠模型均因血管畸形而死亡。从而说明,VEGFR1发挥促进或抑制作用是离不开VEGFR2的,它俩之间相互影响。1.2.2 VEGFR2(KDR或者Flk-1) 大家一致认为,在已发现的几种VEGF受体中,VEGFR2是血管发生和构建的主要调节者,其基因定位于染色体13q12,VEGFR2与配体VEGF结合后介导内皮细胞的增殖、分化等行为,以及血管的形成过程和血管的通透性[5],具体途径可能是VEGF通过细胞膜上的VEGFR2将细胞外的信号传递到胞内,通过PLC--PKC-Raf-MEK-MAPK信号传导通路,将信号传递至核内激活DNA的合成,促进内皮细胞的增殖[6]。与VEGFR1相比,VEGFR-2酪氨酸激酶活性却很强,VEGFR2在细胞内经历了二聚化、配体依赖酪氨酸磷酸化,促有丝分裂、趋化性和促存活信号的功能[7]。1.2.3 VEGFR3(Flt-4) VEGFR3属于酪氨酸激酶家族成员之一,主要表达在胚胎时期的血管内皮细胞和成年期的淋巴管内皮细胞,VEGFC和VEGFD与VEGFR3结合发挥作用,主要可以刺激淋巴管内皮细胞的增殖与迁移,促进淋巴管的新生。但是,在某些病理状态下,VEGFR3也可以发挥促血管生成的作用,也就是说VEGFR3可以表达于肿瘤血管内皮细胞上[8]。近年来许多学者研究发现,VEGFR3在多种肿瘤细胞中也有高表达,且肿瘤组织VEGFR3含量与与淋巴结转移密切相关[9,10]。但是,目前对VEGFR3的信号传导功能了解甚少,也有研究发现只有VEGFR3与VEGFR2共同作用才能发挥对淋巴系统的这种调节作用。有研究表明,在同一组织中VEGFC结合VEGFR3的亲和力比结合VEGFR2的亲和力高3倍多[11]。所以说,VEGFR3对肿瘤新生血管的发育和保持淋巴管完整性非常重要。1.2.4 NRP-1 众所周知,NRP是一种非酪氨酸激酶跨膜蛋白,不能独立转导生物信号,而与VEGF酪氨酸激酶受体形成复合物后才能介导信号传导。NRP-1在几乎所有的肿瘤细胞上均有表达,并主要表达在恶性肿瘤细胞上,但是在相应的正常组织上皮细胞中却没有表达[12],功能丰富,主要在神经发育、血管生成、肿瘤增殖和转移、调节免疫等方面发挥着重要作用,特别是其在肿瘤血管新生及肿瘤转移方面中的作用,越来越受到研究人员的重视[13]。2 肿瘤血管与肿瘤的生长、转移肿瘤的生长及转移的关键步骤是新生血管的形成,大量临床与动物实验都证明,如果没有新生血管供应营养,肿瘤细胞在长到一定程度后不再长大,因此,诱导血管生长将是肿瘤的生长、转移的前提之一。肿瘤的生长大致可以分为两个阶段:第一阶段为血管前期或叫无血管期,此阶段肿瘤处于休眠状态,肿瘤组织体积很小,可在人体内持续数年之久,主要依靠邻近组织血管获取营养,细胞处于稳定更新阶段;第二阶段称为血管期,此期一旦肿瘤组织内某些亚群细胞表型受细胞因子激发,新生血管形成,肿瘤组织获得足够的营养物质和排除代谢废物而迅速增生,即进入快速增长的有血管期。2.1 VEGF与肿瘤血管的形成1971年由Folkman首次提出“肿瘤血管生成”的概念,目前肿瘤血管化的方式有多种,包括芽生式、套叠式、充塞式血管、内皮祖细胞的参与、马赛克血管等多种形式[14]。肿瘤的生长依赖血液不断供给营养,需要新生血管的形成。肿瘤血管生成的机制十分复杂,到目前为止,我们所知道的对血管生成作用较为明确的血管生成因子有VEGF、aFGF、bFGF、PDGF、TGF、TNF、Angiogenin等。而VEGF及其受体是众多血管再生性因子当中的关键因子,能够强烈的刺激内皮细胞进行有丝分裂并最终形成新生血管,也是刺激肿瘤血管生成最强的细胞因子。有许多研究表明,VEGF在肿瘤血管生成中起到关键的作用,如下图1[15]所示能更好的解释VEGF在肿瘤血管生成过程中的不可替代的作用。图1 VEGF在肿瘤血管生成中的作用[15]VEGF促进血管形成的可能机制:VEGF是最强的血管通透因子,作用比组胺强好多倍,可直接作用于血管内皮细胞增加血管通透性,是VEGF重要特性之一,肿瘤血管的高通透性可导致多种血浆纤维蛋白的外漏,为成纤维细胞、内皮细胞等移动侵入提供了暂时的基质成分,促进巨噬细胞、纤维母细胞及内皮细胞生长,从而导致肿瘤血管生成;VEGF诱导降解基底膜过程,促进内皮细胞迁移和生长、诱导血管内皮细胞增殖;内皮细胞管道化分支形成血管环和形成新的基底膜,VEGF还具有促进血管构建等作用。血管直接生成机制仍是公认的肿瘤血管生成的主要机制。也就是说,肿瘤新生血管是在原有的血管基础上延伸扩展而形成的,其过程类似于伤口愈合和胚胎形成过程。研究表明,低氧诱导因子-1(hypoxiainduciblefactor-1,HIF-1)与VEGF存在非常密切的关系,可激活目的基因的转录,VEGF即是其中的靶基因之一,可以导致VEGF-mRNA持续和快速地增加,从而促进肿瘤血管的发生。临床资料表明,大多数恶性肿瘤的血管密集且生长迅速。因此,血管生成在肿瘤的发展转移过程中起到重要作用。众多研究表明,VEGF在各种癌组织中都有过量表达,比如在乳腺癌、卵巢癌、结肠癌、胃癌、鼻咽癌、前列腺癌、黑色素瘤等。有学者认为在前列腺增生、前列腺上皮内瘤变、最后到前列腺癌的演变过程中,VEGF的表达逐渐增高[16],所以VEGF与受体在肿瘤组织的高表达可能对肿瘤的发生、发展有一定的作用,与预后密切相关。2.2
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码









 douglasds
douglasds
本文标题:VEGF及其受体的生物学特性及在肿瘤血管生成中的作用-马莉
链接地址:https://www.777doc.com/doc-4282793 .html