您好,欢迎访问三七文档
当前位置:首页 > 行业资料 > 其它行业文档 > 高温对植物的伤害及耐热性
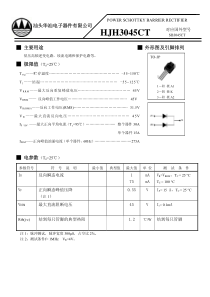
闵雄高温对植物的伤害及耐热机制第一节:高温伤害及植物耐热性概述第二节:高温对植物生理生化过程的影响第三节:高温对植物伤害的生理及分子机制第四节:植物耐热性的生理及分子机制第五节:植物热激蛋白及其分子作用目录高温胁迫引起植物的伤害称热害(heatinjury)。一般是由于高温超过植物生长发育上限温度造成的,主要包括高温害和果树树木日灼伤害两类。第一节:高温伤害及植物耐热性的概述1.高温害•高温害是高温天气对开花至成熟期作物产生的热害,多发生在我国南方早稻和中稻抽穗、开花到成熟期之间,尤其进入盛夏酷热较早的年份。2.果树树木日灼伤•果树日灼是由强烈的太阳辐射增温所引起的果树枝干伤害,也称灼伤。分为夏季日灼和冬季日灼两类。柑橙日灼病冬瓜日灼病冬季涂白的树植物对高温胁迫(hightemperaturestress)的适应和抵抗能力称为耐热性(heatresistance)。3.耐热性温度植株的存活率最低温度最高温度最适温度1.喜冷植物:生长温度为在零上低温(0-20℃),当温度在15-20℃以上即受高温伤害。例如某些藻类、细菌和真菌.二.植物体的温度状况发财树喜冷植物嗜冷细菌2.中生植物:生长温度为10-30℃,超35℃就会受伤。例如水生和阴生的高等植物,地衣和苔藓等。水稻棉花喜温植物3.喜温植物:其中有些植物在45℃以上就受伤害,称为适度喜温植物,例如陆生高等植物,某些隐花植物。有些植物则在65-100℃才受害,称为极度喜温植物,例如蓝绿藻、真菌和细菌等。海栖热袍菌一些作物的临界温度本节小结1.高温对植物的热害可分为:高温害和日灼伤害2.植物对温度状况的分类可分为:(1)喜冷植物(2)中生植物(3)喜温植物3.农作物的临界温度第一节:高温伤害及植物耐热性的概述第二节:高温对植物生理生化过程的影响第三节:高温对植物伤害的生理及分子机制第四节:植物耐热性的生理及分子机制第五节:植物热激蛋白及其分子作用目录第二节:1.高温胁迫对光合作用的影响2.高温胁迫对呼吸作用的影响3.高温胁迫对蒸腾作用的影响4.高温胁迫对植物体内渗透调节物质含量的影响5.高温胁迫对植物体内抗氧化系统的影响6.高温胁迫对植物激素的影响7.高温胁迫对细胞超微结构的影响高温对植物生理生化过程的影响1.高温胁迫对光合作用的影响(1)气孔:叶片是植物进行光合作用的主要器官,在高温胁迫下,光合作用普遍受到抑制。•(2)叶绿素:叶片叶绿素含量变化是叶片生理活性变化的重要指标之一,与光合速率大小具有密切关系。在高温胁迫下,通常呈现出随着时间的延长叶绿素含量下降的趋势,胁迫开始时叶绿素含量下降幅度较小,后期下降的幅度较大(3)光合系统II:PSⅡ位于类囊体膜内侧,是光合电子传递链上第一个多亚基色素蛋白复合体,它能利用光能推动一系列的电子传递反应。在高等植物体内,捕光色素蛋白复合体(LHC)是一类捕获光能,并把能量迅速传递到反应中心引起光化学反应的色素蛋白复合体。PSⅡ的LHCⅡ占光合膜中近50%的色素和大约1/3的蛋白质,生理功能复杂且易被大量提取。高温对PSⅡ影响的主要部位1.天线系统2.放氧复合物3.反应中心(1)高温对PSⅡ天线系统的影响已有实验显示,高温导致豌豆捕光天线构象的变化。Mohanty等研究发现,高温引起豌豆磷酸化的LHCⅡ数量增加,并且磷酸化的LHCⅡ从堆垛区向非堆垛区迁移,高温还会影响与基粒区相连的LHCⅡ的捕光机制。另有实验表明热胁迫导致LHCⅡ从PSⅡ反应中心脱落。但Pospisil和Tyystjarvi用77K发射光谱揭示热胁迫不会引起PSⅡ的LHCⅡ分离。(2)高温对放氧复合体的影响•光合放氧是高等植物和藻类PSⅡ特有的功能,锰原子形成锰簇催化放氧反应,高温处理后锰簇从PSⅡ反应中心复合体中释放,引起PSⅡ放氧活性的丧失。•一旦高温导致PSⅡ放氧复合物失活,光合线性电子传递势必会受到抑制。线性电子传递降低不仅导致光合速率降,而且还可能导致跨类囊体膜质子梯度的降低,而跨类囊体膜质子梯度与植物的光破坏防御机制密切相关放氧复合体中Mn簇的结构,4个锰原子依次连接于PSII的D1蛋白的氨基酸残基上,O和Cl也参与Mn簇的形成。•光合放氧是由锰簇完成的,其根本原因在于锰簇的催化放氧反应,高温处理后锰簇从OEC中释放,导致PSII放氧活性的丧失。组氨酸(3)高温对PSⅡ反应中心复合物的影响一些研究表明高温胁迫可导致PSⅡ反应中心的破坏,抑制激发能从天线色素蛋白到PSⅡ反应中心的传递,并可导致激发能更多的由PSⅡ向PSI的满溢等。Cao和Govinjee提出高温胁迫可导致PSⅡ有活性中心向无活性中心的转化。2.高温对呼吸作用的影响温度对呼吸作用的影响程度取决于呼吸作用的温度系数(Q10)。高温下光呼吸的升高很快(快于暗呼吸的升高),而光呼吸中乙醇酸氧化酶的Q10大于过氧化氢酶的Q10,于是造成H2O2的积累。1、在叶绿体中:RuBP(1,5-二磷酸核酮糖)+O2→PGA(三磷酸甘油酸)+磷酸乙醇酸→乙醇酸2、在过氧化物酶体中:乙醇酸→H2O2+乙醛酸→+谷氨酸→甘氨酸3、在线粒体中:2甘氨酸→丝氨酸+CO23.高温胁迫对蒸腾作用的影响在一定温度范围内,温度升高叶片的气孔开度逐渐增大,加大了水分子从细胞表面蒸发和通过气孔的扩散速率,温度的增加蒸腾速率会加快达到降温的作用,防止叶片被高温灼伤。当温度升高到对植物造成胁迫时,气孔开度开始减小,蒸腾作用随之下降,影响了植物对矿质离子的运输和水分的需求。但时当达到胁迫温度时,气孔关闭,蒸腾能力随之下降,叶温上升,植物正常的生理代谢活动会被扰乱。植物在面对高温胁迫时,出现蒸腾速率下降的情况较多,与植物的耐热性相关。蒸腾作用在植物对水分、矿质离子的吸收、运输和降低叶片温度方面具有重要作用。4.植物体内渗透调节物质的含量渗透调节是植物在胁迫下降低渗透势、抵抗逆境胁迫的一种重要方式,由渗透调节物质来实现。这类物质主要有氨基酸、可溶性糖、可溶性蛋白质、可溶性酚类。在高温环境下,植物主动积累这些物质,抵抗热胁迫的伤害。接下来简单地介绍两种渗透调节物质:(1)脯氨酸(2)可溶性糖植物体内脯氨酸是一种理想的有机溶质渗透调节物,正常情况,植物体内脯氨酸合成酶类有反馈抑制作用,故游离脯氨酸含量不高;当植物处于逆境胁迫下,脯氨酸合成酶类对脯氨酸的反馈抑制的敏感性降低,导致体内游离脯氨酸含量增加。(1)脯氨酸脯氨酸含量的大幅增加有利于水分的保持,从而减轻由于高温引起蒸腾作用加剧而产生的伤害。耐热性不同的两种玉米幼苗在热激过程中都可以诱导Pro的迅速积累,并且随着时间的推移而增加,恢复6h达到最高峰,而后略有下降。另一方面,无论是热激还是恢复过程中,耐热性较强的晴3玉米幼苗Pro积累水平始终高于耐热性较弱的鲁玉13。说明玉米幼苗耐热性的强弱与热激过程中Pro积累水平呈正相关。热激过程中Pro的的水平和耐热性强弱的关系可溶性糖是植物体内一种重要的的渗透调节物质,对植物提高抗性具有重要的作用。一般在高温胁迫下植物体内可溶性糖会累积。大量试验也表明,热胁迫下可溶性糖在细胞质中大量积累,但是在高温下茄子的可溶性糖含量下降,而黄姜的可溶性糖含量表现出低高低趋势。可溶性糖降低的现象可能是植物处在难以承受的温度胁迫下,呼吸作用大于光合作用消耗了贮存的糖类所致。(2)可溶性糖为降低高温造成的伤害,保护酶活性会减轻膜脂过氧化的伤害。CAT和SOD活性在高温胁迫下通常表现为先提高后下降。5.高温胁迫对植物体内抗氧化系统的影响一般认为,尽管SOD可清除O-2,负责催化O-2转变为H2O2,减轻膜脂过氧化对细胞内其他部位的伤害,但这种保护作用是有限的,胁迫后期,当胁迫压力超过植物所能承受的极限时,高温会破坏酶的活性中心,通过改变酶的结构或抑制酶的表达,使得酶活性下降。在高温胁迫下水稻叶片中的SOD活性一直下降。高温胁迫下植物中POD活性变化主要有三种趋势:一是POD活性先升高后下降。在45℃高温胁迫下O-2的迅速产生,使POD活性明显增加,在胁迫3h左右达到高峰,而后迅速下降;并认为随着处理时间的延长,POD的结构可能受到破坏,导致活性下降。二是一些植物中POD活性先下降后上升,三是某些植物经高温胁迫POD活性下降。6.高温胁迫对植物激素的影响植物在生长发育过程中,自身产生一类能调节生长发育的植物激素,在正常环境下这类激素维持在某一恒定的范围内调节着植物的生长。在遭受环境胁迫后植物体内往往迅速积累大量的脱落酸和乙烯,伴随胁迫的解除,它们的含量也回落。胁迫条件下脱落酸(ABA)的积累最为普遍,ABA可以诱导热激蛋白的产生,从而提高植物的抗高温能力,并且ABA含量增加有利于提高植物的抗逆性。7.高温胁迫对细胞超微结构的影响(1).叶绿体(2).线粒体chl:叶绿体M:线粒体GL:基粒片层P:脂质球体CW:细胞壁N:细胞核C:线粒体上的脊高温胁迫处理后的线粒体chl:叶绿体M:线粒体GL:基粒片层P:脂质球体CW:细胞壁N:细胞核C:线粒体上的脊高温胁迫对超微结构影响的结论1.伤害较轻的叶绿体的类囊体片层膨胀,叶绿体体积增大,形状变圆并移向细胞中央;伤害较重的叶绿体的基粒片层膨胀、溶解或空泡化、破裂,基粒垛叠程度下降,脂质球体数量增加、聚合、体积增大;伤害严重的叶绿体的被膜破裂甚至完全解体,基粒外流,仅剩排列紊乱的片层结构,最终导致细胞的不可逆死亡。2.叶绿体形状变圆、体积变大及移向细胞中央,或许是为了减少光线的直接穿透量和受光面积,尽量避免高温灼伤,这是叶绿体对逆境胁迫的生理适应性反应。另外,由于叶绿体类囊体受热降解,降解产物脂质发生集结,故在受伤害较重的叶绿体内部,出现脂质球体数目增多和体积增大现象。本节小结•高温胁迫对植物的生理生化的影响大致可分为7类:7对细胞超微结构的影响1植物光合作用的影响2呼吸作用的影响3对蒸腾作用的影响4对渗透调节物质的影响5对抗氧化系统的影响6对植物激素的影响第一节:植物体的温度状况与热害第二节:高温对植物生理生化过程的影响第三节:高温对植物伤害的生理及分子机制第四节:植物耐热性的生理及分子机制第五节:植物热激蛋白及其分子作用目录•根据高温的强度和植物出现热害的速度和症状,把热害分为三种:•直接伤害,间接伤害和次生水分胁迫伤害第三节:高温对植物伤害的生理及分子机制•直接伤害是指植物在短时间(几秒到几十秒)接触高温后立即表现出来的一种伤害,并可从受热部位向非受热部位传递蔓延。又分为:1.蛋白质变性2.脂类液化•间接伤害间高温引起植物体内一系列变化,这些变化比较缓慢,在高温下经过一段较长的时间才会表现出来。又分为1.代谢性饥饿2.有毒物质积累3.蛋白质合成受阻4.生理活性物质缺乏(1)蛋白质变性1.直接伤害蛋白质是由氨基酸以“脱水缩合”的方式组成的多肽链经过盘曲折叠形成的具有一定空间结构的物质。为什么强调空间结构呢,因为只有形成了空间结构蛋白质才能具有其生物学活性,而高温破坏了蛋白质的空间构型,使蛋白质二级结构和三级结构中起重要作用的氢键因高温断裂,一些维持三级结构的疏水键也遭破坏。这样,蛋白质空间构型被破坏,肽链展开、疏松起来,发生蛋白质变性,失去其原有的生物学特性。高温胁迫对细胞膜稳定性的影响大量研究表明,高温下细胞膜系统的稳定性同抗性成正相关。1.细胞膜损伤导致膜透性增大是高温胁迫的一种普遍现象,膜损伤的分子基础可以是膜蛋变性或膜脂过氧化,也可以是膜结构状态的改变。2.膜蛋白变性或膜脂过氧化与高温胁迫大量活性氧累积导致的氧化损伤有关,而膜结构状态的改变可引起膜脂类流动性的增加或膜脂液化,导致形成非双层脂膜的脂类结构。高温胁迫条件下,处于细胞最外层的细胞膜系统仅次于细胞壁感受到胁迫信号,并通过信号传递途径将胁迫信号传递到细胞内,引起植物体内酶活性的变化,进而影响重要的生理生化反应及逆境诱导基因的表达和调控。但随着胁迫程度的不断增强,胁迫所造成的伤害超过生物体自身的调控能力时,植物就开始表现外观形态的热害症状。由此认为这些脂类变化的主要后果之一是脂类与蛋白质交互作用的紊乱,从而扰乱了膜的正常结构与功能,使质膜的电解质渗透率增加,膜的热
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码







 黛南枫御2
黛南枫御2
本文标题:高温对植物的伤害及耐热性
链接地址:https://www.777doc.com/doc-4477443 .html