您好,欢迎访问三七文档
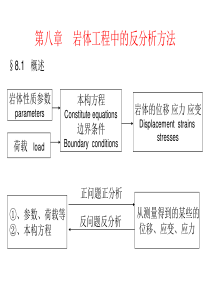
.'.光合作用I:植物对光能的吸收与转换1.叶绿素蛋白是如何进行分类的,它们各有何特点?如果按色素蛋白所含的叶绿素分类,则主要有两类叶绿素蛋白复合体:叶绿素a蛋白复合体和叶绿素a/b蛋白复合体。这些蛋白复合体上还有胡萝卜素或叶黄素,主要起保护光合系统的作用。1叶绿素a蛋白复合体叶绿素a蛋白复合体具有下列的一些共同性质:①结合叶绿素a,含有-胡萝卜素;②为高疏水性蛋白,与反应中心紧密连系;③它们的分子量一般高于叶绿素a/b复合体;④它们都是叶绿体基因编码的蛋白(而a/b复合体通常为核编码)。2叶绿素a/b蛋白复合体叶绿素a/b蛋白复合体又称聚光复合体或捕光色素蛋白复合体(lightharvestingcomplex,简称LHC),它是类囊体膜上最丰富的蛋白复合体,LHC结合了叶绿素a总量的50%和所有的叶绿素b,LHC不参与光化学反应,只起光能传递的作用。叶绿素a/b蛋白复合体也具有一些共同的性质:①结合叶绿素a和b;②分子量在20~30kDa,并且由核基因编码;③光诱导合成,在黑暗中生长的植物缺失a/b复合体,在光下合成;④叶绿素a/b蛋白复合体具有某些共同的氨基酸序列,用免疫方法证明,一种a/b蛋白复合体的抗体可以与其他a/b蛋白复合体有交叉反应,说明它们在结构上有共性,基因序列分析的结果也表明它们具有某些同源的序列。2.类囊体膜上的蛋白复合体的分布有何特点,这样的分布特征与它们的功能有什么联系?蛋白复合体在类囊体上的分布不仅是非均衡的,而且是动态的。蛋白复合体可以沿类囊体膜侧向移动,特别是捕光色素蛋白复合体和细胞色素Cytb6f复合物在基粒类囊体和基质类囊体间的运动在光能的分配和电子的传递的调控中有非常重要的功能。3.在类囊体反应中,涉及哪些蛋白复合体,它们的主要功能是什么?类囊体膜是光合作用中光反应(电子传递和光合磷酸化)的结构基础,光能吸收的捕光色素蛋白复合体、光系统I和光系统II,细胞色素b6f复合体,ATP合成酶。光合电子传递链上的第一个蛋白复合体是光系统II。光系统II是多亚基的蛋白复合体,它利用光能推动一系列的电子传递反应,导致水裂解为氧和质子,释放出氧气,并将电子传到质醌(PQ)。细胞色素b6f复合体在PSII和PSI光反应复合体间进行电子的传递,催化还原态的质醌(PQH2)的氧化和质体蓝(PC)的还原;并与此偶联地把质子从类囊体的基质转移到类囊体腔,建立起跨膜的pH梯度,形成合成ATP的动力;此外,它还有调节PSII和PSI间能量分配以及NADPH和ATP比例的功能。含铜的蛋白质。介于Cytb6/f复合体和PSⅠ之间的电子传递体。铜离子的氧化还原变化传递电子。光系统I从质蓝素接受电子,利用光能推动电子传递,最终将电子传递到铁氧还蛋白。铁氧化蛋白和铁氧化蛋白—NADP+还原酶,严格地说,铁氧还蛋白是光合电子传递的最终产物此外,铁氧还蛋白也参与循环电子传递。再者,铁氧还蛋白是C,N,S同化的中心,在.'.碳同化中,铁氧还蛋白还与硫氧还蛋白的还原有关4.捕光色素蛋白LHCII的结构和功能是什么?LHCⅡ的单体含有12个叶绿素分子(7个叶绿素a与5个叶绿素b),2个类胡萝卜素分子。LHCⅡ的主要功能是进行光能的吸收和传递,提高捕光面积和捕光效率;此外,LHCⅡ在光能分配的调节以及光保护反应中也具有重要的作用。自然状态下有生理活性的LHCⅡ是其三聚体。在三聚体形成过程中,首先形成单体,然后形成三聚体。三聚体的形成与磷脂有关。叶绿素在LHCⅡ中的排列保证了其间的适当距离和角度,使能量可以在其间高效地传递。5.在光合生物中存在哪两类光反应中心,它们有何共同的特点,它们又有何各自特点?高等植物的电子传递链的过程中有两个光反应系统,即PSII和PSI。这两个光反应系统具有一些共同的特点,例如它们核心都是由两条对称的多肽组成,它们都利用“特殊对”的叶绿素分子作为中心色素分子或原初电子供体,它们的原初电子受体是一个叶绿素或去镁叶绿素分子,等等,说明这两个光反应中心可能具有共同的进化来源。这两个光反应中心也有一些完全不同的特点,它们具有不同的最终电子受体,PSII的最终电子受体是质醌,而PSI的最终电子受体是铁氧还蛋白(Ferredoxin)。因此可以将它们分为两类光反应中心:type-I光反应中心和type-II光反应中心。6.如何证明植物中存在两个串联的光反应系统?需氧光合生物具有两个光反应中心的发现最初来自观察到红降和双光增益的现象。Emerson发现当用波长在680nm以上的光照射时,虽然在叶绿素吸收的有效范围内,但光合作用的量子产率却急剧下降,称为红降现象(图6.9)。如果在引起红降的光照(如700nm)的同时,在外加一个短波的光照(如680nm),则量子效率可以提高并且有增益,称之为双光增益(图6.10)。红降和双光增益的现象说明了在光合系统中存在两个相互串联的光反应中心7.光系统II如何进行水的氧化,其中涉及哪些关键装置,氧气的释放步骤如何?光系统II是多亚基的蛋白复合体(图6.13),它利用光能推动一系列的电子传递反应,导致水裂解为氧和质子,释放出氧气,并将电子传到质醌(PQ)。光系统II推动的反应可以表示为:光+PSII2H2O+2PQ——————→O2+2PQH2在水氧化的化学过程中,水被氧化、裂解,放出氧气,需要2分子的水产生1分子的氧气,其中有4个电子的传递,因此PSⅡ需要积累四个氧化等量物以便从两个水分子中提取四个还原等量物(4e/4H+)来产生一个氧气分子8.细胞色素b6f复合体是如何使质子进行跨膜转移的?Q-循环模型认为:在细胞色素b6f上有两个醌的结合部位:一个是还原态醌(QH2)结合和氧化的部位,称为醌的氧化中心(Qo);另一个是氧化态的醌(Q)结合和还原的部位,称为醌的还原中心(Qr)。Qo位点在类囊体膜的囊腔一侧,而Qr位点在类囊体膜的基质一侧。此外,在细胞色素b6f复合体上的两个细胞色素b6有所不同,一个称低电位的b6,或bL,与.'.Qo位点在同侧;另一个称高电位的b6,或bH,与Qr位点在同侧。当QH2在Qo位点被氧化时,第一个电子经QH2→RFe-S→Cytf→PC途径(亦称高电位电子传递途径)进行传递,而它的第二个电子则经低电位电子传递途径传递,即QH2→Cytb(bL)→细胞色素b(bH)。QH2失去两个电子的同时,将两个质子释放到类囊体腔中,被氧化成Q。被还原的bH-把Qr位点的Q还原为半醌阴离子Q-,在下一次QH2电子传递时从基质获取H+,将Q-还原为QH2。这样,每2个QH2在Qo位点氧化就会有4个电子进行传递,2个经高电位电子传递途径将PC还原,另两个传递到Qr位点产生1个QH2,同时,向类囊体腔中释放4个质子,从基质吸收2个质子。这就是一个Q-循环,其总的效果是:氧化1分子QH2为Q,向前传递2个电子,跨膜运转4个质子。9.ATP合成酶是如何进行ATP的合成的?化学渗透学说认为:电子传递链的各组分在产生ATP的膜系统中的分布是不对称的,电子在电子传递体的传递过程起着质子泵的作用,因而在膜的两侧形成了pH梯度(ΔpH)及电位梯度。在这个梯度的驱动下,质子穿过内膜上的ATP酶复合物流回到基质,其能量促使ADP和Pi合成ATP。变构学说认为:在ATP形成过程中,与ATP合成酶活性密切有关的F0的3个亚基各具一定的构象,分别称为紧张(tight)、松弛(loose)和开放(open),各自对应于底物的结合、产物形成和释放等三个过程。构象的相互依次转化是和质子的通过引起亚基的旋转相偶联的。当质子顺质子电化学梯度流过F0,使亚基转动,亚基的转动引起亚基的构象依紧张→松弛→开放的顺序发生改变,使ATP得以合成并从催化复合体上释放。具体说,ADP与Pi与开放状态的亚基结合;在质子流的推动下亚基的转动使亚基转变为松弛状态并在较少的能量变化情况下,ADP与Pi自发地形成ATP,再进一步转变为紧张状态;亚基继续变构成松弛状态,使ATP被释放,并可以再一此结合ADP与Pi进行下一轮的ATP合成。在ATP的整个合成过程中,主要耗能的步骤是ATP的释放,而非ATP的合成。10.什么是光能分配的状态I和状态II,植物是如何实现光能在两个光系统间的平衡分配的?完整叶绿体膜在光下与Mg2+、ATP保温可诱导捕光色素蛋白LHCⅡ磷酸化,同时使吸收的激发能有利于向PSI分配,即诱导状态Ⅱ;而在暗中这一过程可逆转,已磷酸化的LHCⅡ发生脱磷酸化,同时激发能有利于向PSII分配,即诱导状态Ⅰ。天线移动假说认为:捕光的天线蛋白LHCⅡ磷酸化后,会从PSII分布的基粒类囊体的垛叠区向PSI分布的基质类囊体移动,因此扩大了PSI的捕光面积,使吸收的光能更多的向PSI分配;反之,当LHCⅡ脱磷酸化后,则从基质类囊体向基粒类囊体垛叠区移动,其结果是扩大了PSII的捕光面积,使吸收的光能更多的向PSII分配11.什么是光抑制,植物体进行光保护的机制有哪些?光抑制(photoinhibition)是指多余光能对光合作用产生抑制作用,使光合作用的量子效率下降的现象。在光合生物在进化的过程中,产生了多层次的光保护机制首先,植物通过各种方式减少光能的吸收,以达到降低光破坏的目的。叶片是光能吸收的主要器官,减少叶面积,在叶表面形成叶毛或表面物质,改变叶与光的角度等都可以降低光能的吸收。例如在高光照的地区,植物叶片常较小;在干燥、高光照的沙漠地区,一些植物的叶变态为刺(当然这和水分平衡也是有关的)。一些植物叶的表面形成叶毛结构,形成角质或腊质层不仅可以减少水分的散失,而且也可以减少光的吸收。.'.植物细胞中有过氧化氢酶、过氧化歧化酶、抗坏血酸还原酶、谷胱甘肽还原酶、抗坏血酸氧化酶等酶系统,可以使氧自由基、过氧化物等失活,防止对植物细胞的破坏。植物中的酚类化合物、类胡萝卜素等可以直接淬灭单线态氧。当叶绿素吸收光能较多时,非光化学淬灭就会增加而使多余的激发能通过其他途径被消耗掉。光合作用(II):光合碳同化1.卡尔文循环中的主要酶促反应及其生理意义是什么?图7-3核酮糖-1,5-二磷酸羧化酶催化的羧化反应2.说明rubisco的特点及其对光合作用的重要性。核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)具有双重功能:既能使RuBP与CO2起羧化反应,推动C3碳循环;又能使RuBP与O2起加氧反应而引起C2氧化循环即光呼吸。3.叶绿体类囊体膜内pH值和Mg2+浓度在光暗条件下如何变化?对光合碳循环的意义?Rubisco被CO2与Mg2+活化,叶绿体间质中H+和Mg2+浓度有光暗的变化,光驱动H+从间质到类囊体的囊内腔,间质中pH从7变化到8左右,这是rubisco催化反应的最适pH。伴随H+进入囊内的是Mg2+从囊内腔到间质中,Rubisco与Mg2+结合而活化。光下,rubisco活化酶经硫氧还蛋白活化后水解ATP,使rubisco构象变化、释放RuBP后才能通过结合CO2与Mg2+而活化(图7-4)。CO2结合到rubisco大亚基活性位点的赖氨酸的氨基上,形成氨基甲酸酯Lys-NH-CO2,这样Mg2+再迅速结合到上面去,此时酶构象发生变化而具催化功能(图7-5)。然后rubisco才能依次结合RuBP和CO2,催化羧化反应。4.比较光合作用碳代谢的C3途径和C4途径。C3途径C4途径场所叶绿体基质羧化叶肉细胞细胞质固定维管束鞘细CO2受体RuBPPEP关键酶RubiscoPEPC和Rubisco最初产物PGOA★CO2-★CO2-★CO2?上部??下部?RuBP不稳定的酶结合中间物3-磷酸甘油酸★CO2-★CO2-★CO2?上部??下部RuBP不稳定的酶结合中间物3-磷酸甘油酸.'.5.说明光对光合酶的调节及其意义。虽然CO2同化不直接需要光,但由于需要的同化力来自光反应,而且重要的酶都是光活化的,所以在暗中,由于缺乏同化力ATP与NADPH,催化固定CO2的酶不活跃,加之气孔关闭,CO2减少,因而光合作用、包括CO2同化的卡尔文循环不能继续进行。6.光呼吸有什么特点?说明C2循环和C3循环的关系。在光呼吸碳
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码








 justis136
justis136
本文标题:植物生理练习题
链接地址:https://www.777doc.com/doc-6141368 .html