您好,欢迎访问三七文档
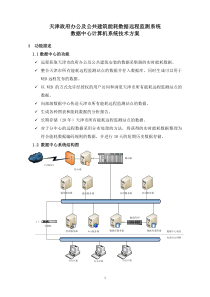
1第八章免疫应答(Immuneresponse)第一部分B细胞介导的体液免疫第一节免疫应答的概念抗原性物质进入机体后激发免疫细胞活化、分化和效应过程称之为免疫应答(Immuneresponse,IR),在高等动物和人体内存在有结构复杂的免疫系统,是由免疫器官、免疫细胞和免疫分子组成的,免疫应答是由多细胞系完成的。免疫应答过程的效应表现,主要是以B细胞介导的体液免疫和以T细胞介导的细胞免疫。这二种免疫应答的产生都是由多细胞系完成的,即由单核吞噬细胞,T细胞和B细胞来完成的,这一过程可包括:(1)免疫细胞对抗原分子的识别过程(抗原分子与免疫细胞间的相互作用)(2)免疫细胞的活化和分化过程(免疫细胞间的相互作用)(3)效应细胞和效应分子的排异作用1.抗原识别阶段此阶段可包括对抗原的摄取、处理加工、抗原的呈递和对抗原的识别分别由MΦ,T和B细胞完成。2.免疫细胞的活化和分化阶段此阶段可包括抗原识别细胞膜受体的交联,膜信号的产生与传递,细胞增殖与分化以及生物活性介质的合成与释放,主要由T和B细胞完成。3.免疫应答的效应阶段此阶段主要包括效应分子(体液免疫)和效应细胞(细胞免疫)对非已细胞或分子的清除作用,即排异效应,及其对免疫应答的调节作用。在此阶段除抗体和效应T细胞参预外,免疫增强系统也参预参加才能完成排异和免疫调节作用。2免疫应答过程识别活化与分化效应抗原的摄取处理和加工抗原的呈递抗原识别膜受体的交联膜信号的产生与传递细胞增殖与分化生物活性介质的合成与释放对靶细胞或分子的排异作用引起组织的损伤作用免疫应答的调节作用MΦ抗原TBT与B细胞的增殖与分化抗体的产生与释放细胞因子的产生与释放效应T细胞的产生免疫记忆细胞的产生抗体分子免疫保护抗感染抗肿瘤效应T细胞免疫病理自身免疫变态反应移植排斥抗宿主反应免疫增强系统补体分子细胞因子K/NK细胞MΦ粒细胞肥大细胞红细胞血小板3第二节抗体产生的细胞学基础一、抗体产生:当第一次用适量抗原给动物免疫,需经一定潜伏期才能在回液中出现抗体,含量低,且维持时间短,很快下降,称这种现象为初次免疫应答。若在抗体下降期再次给以相同抗原免疫时,则发现抗体的潜伏期较初次应答,TD抗原可引起再次应答,TDI抗原只能引起初次应答。初次应答主要是IgM,抗原结合力低,再次应答则主要为IgG,为高亲合性抗体。二、免疫细胞在抗体生成中的作用:抗体产生需要三种细胞参预,即单核吞噬细胞系,T细胞和B细胞分,抗体产生不只是涉及抗体与免疫细胞间的相互作用(对抗原的识别和抗原的激发作用),同时也涉及免疫细胞间的相互作用(免疫细胞活化,增殖与分化),MΦ是抗原处理和呈递细胞,无特异识别抗原的功能。T细胞系主要是TH和TS,它们对免疫应答有调节功能,所以是免疫调节细胞,有特异识别抗原的功能,B细胞系是既具有呈递抗原的作用又是产生抗体的细胞,也具有特异识别抗原的功能。4(一)MΦ的作用MΦ在免疫应答的全过程都发挥重要作用,在抗原识别过程中,即在免疫应答的透导期,它表现为具有摄取,处理加工,存贮和呈递抗原的作用。它活化后还能分泌多种细胞因子,其合成和分泌的IL-1有促进T和B细胞的活化作用。抗原物质进入体内后,必须先经MΦ摄取,加工处理,然后才能呈递给淋巴细胞,MΦ细胞表面有多种受体分子,但无抗原识别受体,它主要以吞噬、吞饮和被动吸附等方式捕捉抗原,可摄取任何抗原性物质,为非特异性摄取。(二)淋巴细胞的作用淋巴细胞具有抗原识别受体,所以T和B细胞都是抗原识别细胞。B细胞表面抗原受体是膜Ig分子,它可以识别天然蛋白质抗原分子表面的构像抗原决定簇,在识别抗原时无MHC限制性。T细胞抗原识别受体异二聚体分子,即TcRαβ,它能同时识别经加工处理的序列决定簇肽片段和MHC分子,所以有MHC限制性。三、细胞因子在抗体产生中的作用1.细胞因子的作用既无抗原特异性,也无MHC限制性。2.在B细胞产生免疫应答的不同时期有不同的细胞因子起作用。3.细胞因子还可作用于路(Bystandcr)B细胞使之活化。4.在刺激B细胞增殖分化的细胞因子中,有些是来源于巨噬细胞或其他非T细胞所产生。第三节抗体形成过程中免疫细胞间相互作用一、巨噬细胞与TH细胞之间的相互作用TH细胞必须由静止状态的TH转变活化才能辅助B细胞产生抗体。近年的实验证明,在抗原呈递细胞(APC)表面至少有二种分子与TH细胞的活化相关。一种是抗原呈递分子,它是由MHC分子组成,它可与外源性或内源性抗原肽片段结合,然后运送至细胞表面并呈递给T细胞,通过TCR/CD3刺激产生第1活化信号。另一种分子即所谓协同刺激分子(costimulatingmolecules,CM)它是由一组粘附分子组5成,这能促进APC与T细胞的直接接触,而且诱导信号传递的功能,这组分子可与T细胞上的协同刺激分子受体(costimulatorgmoleculesreceptor,CMR)结合,刺激其产生协同刺激信号,即第2信号。在这二种信号的作用下,才能使T细胞活化并合成和分泌IL-2和表达IL-2R最终导致细胞分裂和克隆扩增。如无第2信号存在则T细胞不被活化也不引起克隆扩增,处于克隆不应答状态(clonolanergy)。阻断或给予第2信号,可以人为调节免疫应答,使之增强或抑制,对免疫治疗提供了新的手段。二、TH细胞与B细胞之间的相互作用1.B细胞对T依赖抗原的应答当大量抗原进入未经免疫的机体后,诱发初次免疫应答时其抗原呈递细胞多由巨噬细胞完成。经MΦ活化TH细胞后再由活化的TH细胞辅助B细胞产生抗体和形成记忆B细胞。但当再次免疫应答发生时,抗原呈递细胞则主要由已扩增的B细胞克隆承担。由于其膜Ig受体亲和力增高故对少量抗原也能摄取,故可取代巨噬细胞的抗原呈递作用。B细胞与MΦ相同,也是通过抗原呈递作用与TH细胞相互作用,即B细胞可通过其膜Ig受体与蛋白质抗原,即T依赖性抗原结合,经内化作用将抗原摄入。然后经过加工处理,使T细胞决定簇与MHC-II类分子结合,形成MHC-肽分子复合物并运送至B细胞表面,将其呈递给TH细胞,所以B细胞与TH细胞间相互作用也是MHC限制性的。B细胞和TH细胞通过其粘附分子及抗原呈递作用,可彼此直接接触,并能相互诱导使之活化。活化B细胞在TH细胞的辅助下,B细胞最终增殖分化为合成和分泌各类免疫球蛋白分子的浆细胞。62.B细胞对非T细胞依赖抗原的应答B细胞对非T细胞依赖抗原(T1)的活化机制与T依赖抗原完全不同。对I型T1类抗原,决定簇(细菌脂多糖抗原)在高浓度下,是B细胞多克隆活化剂,它与B细胞表面抗原受体结合,具有促分裂素(mitogen)的结构,可与其相应受体结合,使B细胞多克隆活化。而II型T1抗原的结构特点是具有多个重复出现的抗原决定簇呈线性排列在体内不易降解(如肺炎球菌多糖抗原),这些抗原对B细胞抗原受体亲和力强,由于受体交联而使B细胞活化。T1抗原虽可被APC摄取,但不能被加工处理,所以不能与MHC-II类分子结合,故B细胞对T1抗原的应答无TH细胞参预,只表现初次应答的特性而不出现再次应答的一系列变化。3.巨噬细胞与B细胞的相互作用7第四节免疫记忆用同一抗原再次免疫时,可引起比初次免疫更强的抗体产生,称之为再次免疫应答或免疫记忆,无论在体液免疫或细胞免疫均可发生免疫记忆现象。在体液免疫时,对TD抗原的再次应答可表现为抗体滴度明显上升,免疫球蛋白类别可由IgM转换为IgG,而且抗体亲和力增强。1.免疫记忆细胞:在免疫应答过程中,既能产生B记忆细胞(BM),也能产生TH记忆细胞(THM)免疫记忆现象可以解释为对特异抗原,应答的淋巴细胞数量增加的现象。2.免疫球蛋白类别的转换:在初次应答时开始出现的抗体是IgM,当迟到高峰时才开始出现IgG,而IgG高峰虽出现晚,但能维持较长时间。在再次应答时产生IgG的潜伏期明显缩短,水平更高。这种IgM转换为IgG只是Ig分子的类别变化,其识别抗原的特异性其仍相同。这种转换可能由产生IgM的细胞变为产生IgG的细胞,而不是由不同亚群的B细胞产生的。3.抗体亲和力的变化在抗体生成过程中,抗体分子的平均亲和力随时间的延长而增加,这种现象称为抗体分子亲和力的成熟。初期免疫应答可因存在较大量的游离抗原分子,因此,与低亲和力受体B细胞结合较多。当抗原量逐渐减少时,则与带有高亲和力受体细胞的结合多于低亲和力受体细胞。因之抗体分子的平均亲和力随时间延长而增高。8第五节体液免疫的效应免疫应答最终效应是将侵入机体的非已细胞或分子加以清除,即排异效应。但抗体分子本身只具有识别作用,并不具有杀伤或排异作用,因此,体液免疫的最终效应必须借助机体的其它免疫细胞或分子的协同作用才能达到排异的效果。1.抗体分子的中和作用(结合侵入机体的病毒,外毒素分子)。2.抗体分子的调理作用(与抗原形成的复合物、促进吞噬细胞的排除功能)。3.补体介导的细胞溶解作用(补体分子可经第一活化途径工旁路途径溶解靶细胞)。4.抗体依赖细胞介导的细胞毒性作用。(参预抗体依赖细胞介导的细胞毒作用ADCC,如巨噬细胞,中性粒细胞和杀伤细胞)第六节体液免疫应答的调节1.抗体的反馈调节与抗原结合,并被清除。2.免疫抑制细胞作用当免疫应答发展到一定程度时,即能透发抑制性T细胞,分泌特异性抑制因子(TSF),和它参预的网络调节。3.免疫网络调节(immnenetwork)促使抗独特型抗体的产生,促进和抑制免疫应答。9第二部分T细胞介导的细胞免疫第一节细胞免疫的概率凡是由免疫细胞发挥效应以清除异物的作用称为细胞免疫。参预的细胞称为免疫效应细胞。目前认为具有天然杀伤细胞(NK)和抗体依赖的细胞介导的细胞毒性细胞(antibodydependentceel-mediatedcytotoxicity,ADCC),如巨噬细胞(MΦ)和杀伤细胞(K)以及由T细胞介导的细胞免疫均属细胞免疫的范畴。前二类免疫细胞在其细胞表面不具有抗原识别受体,因此它们的活化无需经抗原激发即能发挥效应细胞的作用,故可视之为非特异性细胞免疫。而效应T细胞则具有抗原识别受体,因此它们必需经抗原激发才能活化,发挥其效应细胞的作用,故可视之为特异性细胞免疫。由T细胞介导的细胞免疫有二种基本形式,它们分别由二类不同的T细胞亚类参预。一种是迟发型超敏性T细胞(TOH,CD4+),该细胞和抗原起反应后可分泌细胞因子。这些细胞因子再吸引和活化巨噬细胞和其它类型的细胞在反应部位聚集,成为组织慢性炎症的非特异效应细胞。另一种是细胞毒性T细胞(TC,CD8+),对靶细胞有特异杀伤作用。引起细胞免疫的抗原多为T细胞依赖抗原(To抗原),与体液免疫相同,参预特异细胞免疫的细胞也是由多细胞系完成的。即由抗原呈递细胞(巨噬细胞或树突状细胞)、免疫调节细胞(TH和TS)以及效应T细胞(TDTH和TC)。由T细胞介导的主要细胞免疫现象有:(1)迟发型超敏性反应;(2)对胞内寄生物的抗感染作用;(3)抗肿瘤免疫;(4)同种移植排斥反应;(5)移植物抗宿主反应;(6)某些药物过敏症;(7)某些自身免疫病。10第二节CD4+T细胞介导的细胞免疫一、CD4+T细胞在DTH反应中的作用这是由CD4+T细胞激发的特异性的细胞免疫应答,它可引起组织的慢性炎症,它是以淋巴细胞(主要是T细胞)和单核吞噬细胞系细胞浸润为主的渗出性炎症。由于免疫细胞的激活,增殖和分化以及其它炎症细胞的聚集需要较长时间,所以炎症反应发生较迟,持续时间也长,故称此种炎症反应为迟发型,超敏反应(delayedtypehypersensitivty,DTH),诱发这种反应的T细胞称为迟发型超敏性T细胞(TDTH)。其后在小鼠体内证明参预这种反应的淋巴细胞是Lyt-1+细胞(相当于人的CD4+T细胞),其表型与辅助性T细胞(TH)相同,并且两者在识别抗原的MHC限制性上也相同。但两者在功能上完全不同,前者介导细胞免疫,而后者参预体液免疫,再据其合成和分泌的细胞因子不同,可分为TH1和TH2两种亚类。TH1介导细胞免疫,TH2参预体液免疫。二、CD4+T细胞的活化CD4+T细胞的活化需有抗原呈递细胞参预,主要为巨噬细胞(MΦ)
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码




![米花岭隧道机械化施工设备配套[1]](/doc-136068.png)



 也同
也同
本文标题:第八章__免疫应答
链接地址:https://www.777doc.com/doc-2085980 .html