您好,欢迎访问三七文档
当前位置:首页 > 行业资料 > 酒店餐饮 > 肠道菌群与粘膜免疫系统
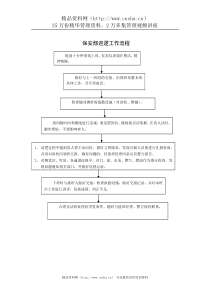
肠道菌群与粘膜免疫系统MichaelH.Chapman,IanR.Sanderson英国伦敦大学Barts&TheLondon,圣玛丽医院成人及儿童胃肠病科,TurnerStreet,伦敦E12AD,英国前言出生时胃肠道是无菌的,但很快有种类繁多的细菌定植,因此成为人体接触病原微生物的首要部位,甚至90%的微生物是通过胃肠道进入人体的。胃肠道最主要的功能在于摄取营养和维持体液的平衡以驱除有害的微生物和其它一些毒素物质。我们就胃肠道粘膜免疫系统的基本组成及病原微生物如何与其和肠道功能的其它方面相互作用进行综述。肠道的正常菌丛出生时胃肠道的粘膜免疫系统的活性较低,与成年人比较淋巴细胞和Payer斑都较少。出生后经口菌群定植很快发生。肠道菌群在不断地发生变化直到成年才变得稳定,且会随着饮食结构的改变而发生变化。例如,母乳中IgA水平在婴儿期就起着非常重要的作用。胃肠道的菌群总量是非常大的,近50%的粪便是细菌,约为1012/克。随着胃肠道的长度发生变化,其细菌数目和种类也不同。除口腔外,菌落随着胃肠道的延伸而逐渐增多,而胃和近端小肠却只有少量的以革兰氏阳性为主的细菌。菌群在小肠远端和结肠变成一个非常复杂的微生物环境。这些区域也正是炎性肠疾病(IBD)最容易受累的部位,这使我们推测粘膜免疫系统对胃肠道菌群的无效或不正常的反应在这些疾病的发病机制中扮演了非常重要的角色。胃肠道的菌群总量是非常大的,粪便中近50%是细菌,约为1012/克粪便由于许多方面的原因定义正常的肠道菌群是非常困难的。已知有超过500种不同种类的微生菌群在肠道定植,在回肠末端及结肠部的主要定植菌群包括乳酸杆菌、双歧杆菌、肠球菌和拟杆菌[1-2]。由于许多菌群无法在体外进行培养因而对其研究也一度受到阻碍,近来,借助于新的研究方法如变性梯度凝胶电泳(DGGE)和荧光原位杂交(FISH,利用菌群特异性探针对其进行组织定位)使对这些菌群研究取得重大进展。肠腔和其相关联的粘膜上微生物菌群的数量和类型也是有差别的[3]。粘膜相关菌群很可能与肠道正常功能和疾病有关,但由于研究粘膜相关菌群技术上的复杂性,绝大多数研究集中于对粪便菌群的分析。正常菌群的作用是复杂的,具有对抗病原微生物的入侵,在宿主的营养中起着至关重要的作用。这些菌群被定为共生菌群,忽视了其在维持宿主健康方面的积极作用。对宿主最主要的营养益处包括维生素的合成和分解复杂多糖产生短链脂肪酸[4],这些作用单靠肠道组织本身无法完成而有赖于肠道菌群的作用。无菌环境中成长的动物为了达到与正常菌群定植的动物的同样的体重其对营养成分的需求就要多得多。肠道粘膜免疫系统的功能组分肠道粘膜具有对抗潜在病原微生物入侵的生理屏障和免疫防线的双重作用,下面我们就肠道防御系统的功能组成和他们遭破坏时病理性变化举例说明,接着讨论一些肠道和定植于其上的微生菌群相互作用有益的方面。胃肠道粘膜具有对抗潜在病原微生物入侵的生理屏障和免疫防线的双重作用屏障功能胃肠道具有大约400m2的巨大表面积以完成其营养物质吸收的这一首要功能。这么大的面积需要保护不被病原微生物的入侵。肠道上皮层作为高度选择性的吸收表面同时也成为微生物入侵时非常有效的屏障,其屏障功能的机制如图1所示。消化液中的胃酸和蛋白水解酶破坏了大部分进入肠道的病原微生物。幽门螺杆菌是少有的几个例外的一种,这种细菌可以通过改变其周围的pH而在这种酸性环境生存下来。上皮细胞间形成有效的紧密连接,成为防止微生物入侵的直接屏障。屏障破坏是绕过淋巴滤泡正常调节作用最关键的一步,并由此导致一些过度的免疫反应如炎性肠病(IBD)等。通过甘露醇通透试验证实肠道菌群组成本身的改变对肠壁的通透性具有直接的影响[5]。由杯状细胞分泌的粘液层可破坏微生物的定植和增加其清除。粘液层糖蛋白竞争肠上皮细胞上结合位点从而抑制细菌入侵,杯状细胞分泌产生的粘液量和组成的不同取决于肠腔菌群的局部信号和肠腔菌群本身[6]。乳酸杆菌可降低其他肠道微生物菌群的转位,这一机制涉及上皮细胞产生的粘蛋白的上调[7]。每24-96小时就更新一次的肠上皮细胞减少了肠腔微生物入侵的时间,并且能够快速修复由于炎症或创伤引起的粘膜破坏[8]。最后,病原菌和“正常的”共生菌之间存在着生存空间与营养物质的竞争,例如定植抵抗作用,这种现象最明显的例子就是抗生素应用导致正常结肠菌群的破坏从而使艰难梭菌生长诱导伪膜性肠炎的发生。一种生物在粘膜表面存活(而不是肠腔)依赖于其在通过以上提到的机制被清除之前他们的分裂和侵袭能力,能够穿透这一屏障的微生物还将受到以下谈到的各种粘膜免疫系统组分的调控。先天性免疫尽管存在肠道的屏障功能,仍然有稳定数量的细菌通过肠道粘膜,但他们被吞噬细胞活性所控制,这项功能主要有中性粒细胞和巨噬细胞通过识别多种细菌的共同抗原如脂多糖(LPS)而实现的。识别非特异的微生物抗原是通过类型识别受体如Toll样受体家族实现的,后者也在补体系统活化、细胞因子反应和抗原递呈方面起作用[9]。肠上皮Paneth细胞(潘氏细胞)可分泌强有力的抗微生物多肽,其中包括防御素和cathelicidies,这些成分形成了对所有多细胞微生物共有的古老的防御机制,并在所有的上皮表面产生。细菌及其产物诸如LPS可直接刺激防御素和其它肽类的释放,这些肽类物质具有多方面的作用,其中包括吞噬趋化作用和破坏细菌细胞壁直接杀伤微生物[10]。免疫缺陷病人或动物的自然史对于我们理解免疫系统不同组分的重要性提供了很好的注解。非特异性免疫系统缺陷病人(如中性粒细胞/巨噬细胞功能不全)往往罹患最严重疾病,例如频繁发生由肠道感染引起的脓毒血症非常的普遍。巨噬细胞杀伤活性缺陷动物会很快死于由共生的肠道微生物引起的脓毒血症[11]。IgA缺陷病人引起的疾病较轻微而不易在临床上检测到。这些发现证实先天性免疫反应在保护宿主中的重要性。适应性免疫完整的先天性免疫反应很少引起微生物穿透粘膜屏障导致严重损害,那么为什么肠道内有这种明显的“生理性的炎症反应”状态呢?另外一个与此相关联的关键问题是粘膜免疫系统如何识别不同的肠腔病原微生物并与其反应?膜免疫系统必须能够抑制对饮食中抗原和共同菌群的过激反应,同时产生对病原微生物的有效反应机制。适应性免疫反应很可能在区别以上的反应中起重要作用,调控这种反应机制的破坏是炎症性肠病和坏死性小肠结肠炎等疾病的可能机制之一。下面就适应性免疫系统的组成予以阐述。IgAIgA通过结合、中和和防止病原微生物黏附于上皮而防御特异性抗原的入侵。它是由固有层的胞浆细胞产生,通过上皮转运、分泌到肠腔与其他分泌组分一起形成分泌型IgA(SIgA),与传统的抗体由T细胞辅助系统免疫应答中产生不同,肠粘膜上的SIgA产生似乎不依赖于T细胞[12],是由来源于胸膜腹膜的B1细胞产生,而与T细胞辅助的骨髓源的B2细胞不同。肠上皮细胞除了吸收和屏障功能外,肠上皮细胞在粘膜免疫反应中还扮演着重要的整合作用。其中最值得注意的是覆盖于Peyer’s斑和淋巴样聚集的特异性M细胞(集合淋巴小结),将在以后章节叙及。即便是非特异的上皮细胞亦能通过表达细胞因子受体和表达递呈抗原的主要组织相容性复合物(MCH)I/II类分子来显示其免疫活性和免疫反应性,他们亦可合成细胞因子、一氧化氮和防御素[13,14]。上皮细胞活性的激发源于与微生物的直接接触或通过粘膜细胞层淋巴细胞的活化。上皮细胞也在肠腔抗原/微生物菌群和粘膜淋巴结的交互作用的信号传导中起部分作用,其中涉及上调细胞表面受体如CD86和ICAM-1[15]。上皮细胞出现特异类型的受体表达和细胞因子释放表明上皮细胞接触到不同的共生菌群时,对临近的上皮内淋巴细胞产生特异不同的免疫信号通道[16],一些研究人员的结果表明肠道上皮细胞的抑制功能不全在炎症性肠病的发病中起重要作用[17]。粘膜相关淋巴组织(MALT)肠粘膜上有数量众多的淋巴细胞,仅从他们的绝对数目就可充分提供肠道的所谓“生理性炎症”状态的证据。这些淋巴细胞可在整个粘膜和粘膜下层广泛分布也可作为次级滤泡不连续的聚集分布。粘膜免疫系统必须能够抑制对饮食中抗原和共同菌群的过激反应,而对病原微生物产生有效反应。适应性免疫反应很可能在区别以上的反应中起重要作用。次级淋巴滤泡具有和淋巴结同样的结构和功能。最熟知的次级滤泡是小肠Peyer氏斑,位于固有层中特殊的上皮细胞-M细胞的下面(图2)。M细胞的主要作用似乎在于吸收和转运肠腔抗原到Peyer氏斑的淋巴细胞,抗原传递可以是以小分子物质或吞噬的整个病原体的形式。许多微生物如沙门氏菌、霍乱弧菌[18]和脊髓灰质炎病毒[19]都误用这一有效的细胞内摄作用作为进入肠道粘膜的途经。结肠次级滤泡组织没有Peyer氏斑那么有组织但有同样的功能,其下面的M细胞有识别肠腔抗原作用[20]。Peyer氏斑和其它次级滤泡是适应性免疫应答的主要作用部位。观察性研究发现在无菌环境中出生和饲养的动物其Peryer斑较小,同时伴有CD4+、CD8+和IgA分泌型淋巴细胞数目的减少。细菌植入后,可导致淋巴细胞的快速增殖和Peyer氏斑的增大。生发中心的反应在感染后14天达高峰之后开始减弱。当第一次接触共生菌群时,有相当多的菌群穿过粘膜且可从肠系膜淋巴结中或脾脏中培养出这些菌群。随着适应性免疫反应的激活,其免疫活性可自限性的升高从而维持正常微生物菌群局限于肠腔内[20]。次级淋巴滤泡中粘膜相关淋巴组织(MALT)的激活的经典模式表明:抗原激活的B细胞和T细胞的聚集发生在Peyer氏斑中位居圆顶区的巨噬细胞和树突状细胞抗原递呈之后,这种活化主要是IL-12引起的Th1类型反应[21]。然后在此淋巴滤泡内发生B、T细胞克隆性的扩展。与数量有限的高度保守的外源性抗原(如LPS)能被先天性免疫应答细胞识别相反,淋巴细胞对大量的特异性抗原能生成高度选择性的受体。IgA分泌型B细胞和活化的T细胞(主要是CD45RO记忆细胞)在肠系膜淋巴结和血中循环,之后才定居肠粘膜并成为粘膜固有层(LPLS)和上皮间淋巴细胞(IELS)。最初的定居信号是MADCAM-1,它可在粘膜组织的血管内皮细胞中表达,需要有淋巴细胞的活化。这样,由肠道局部病原触发的免疫反应可以在整个肠粘膜的防御中起作用。通过T、B淋巴细胞恰当的克隆选择或清除过程,成熟的淋巴细胞进入外周血循环中。对病原微生物选择性的反应和对共生微生物及食物抗原的相对耐受机制还不清楚。我们知道,若没有源于抗原递呈细胞(APC)的共同刺激信号的作用,识别食物抗原淋巴细胞可以凋亡或变得无活性(无变应性)。加工刺激外源性抗原的抗原递呈细胞激活了多条细胞内通路如淋巴细胞共同激活的B7细胞。源于抗原递呈细胞的局部细胞因子的活性在确定未成熟Th0细胞的最终表型中也发挥作用。总之,以IL-10为主的刺激将会产生调节T-细胞(T-regCells)。IL-4促使Th2细胞分化而IL-12促进Th1细胞分化。对乳酸杆菌的研究发现甚至不同的乳酸杆菌菌株通过改变树突状细胞功能和细胞因子的释放就能刺激不同的免疫反应[22]。阑尾组织中有密集的淋巴滤泡,但其功能至今还不清楚。许多研究表明在人体它是没有功能的,然而,有人观察到行阑尾切除术的病人其溃疡性结肠炎的发病率下降或比有阑尾的病人溃疡性结肠炎的发病时间延迟,程度较轻。这一观察结果表明阑尾具有尚未证实的免疫功能[23]。固有层淋巴细胞(LPLs)和上皮内淋巴细胞(IELs)固有层淋巴细胞和上皮内淋巴细胞代表了免疫系统的效应细胞。一旦淋巴滤泡中的T或B淋巴细胞激活,他们就能通过淋巴系统进入到外周循环,然后需要淋巴细胞的活化归巢到粘膜区域[24]。这些部位可以是肠道粘膜的任何地方,主要取决于适当的受体表达、化学因子和其他细胞活化信号,而与他们首次被激活的部位没有必然的联系。固有层淋巴细胞(LPLs)固有层淋巴细胞中CD4+和CD8+T细胞与外周血淋巴细胞的比例是相似的,即CD4+/CD8+=2:1,CD4+主要呈现活化的形式,他们表达CD45RO标记;CD8+细胞表达CD28,表明其主要有细胞毒功能[25],肠粘膜活化T细胞比外周血中更高是由于
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码









 surprised
surprised
本文标题:肠道菌群与粘膜免疫系统
链接地址:https://www.777doc.com/doc-5237428 .html