您好,欢迎访问三七文档
当前位置:首页 > 电子/通信 > 综合/其它 > 高级植物生理复习.总结
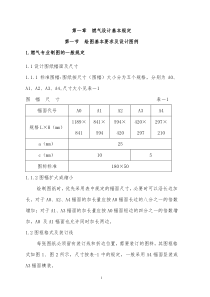
一、生物强化铁锌生物强化的途径及生物学依据。机理Ⅰ植物吸铁机制双子叶植物和非禾本科单子叶植物铁吸收模式:两步过程:第一步包括铁缺乏导致产生三价铁还原酶,该酶用来还原胞外Fe(Ⅲ)螯合物,释放出游离的Fe2+离子。第二步是Fe2+通过特定的Fe2+转运蛋白或二价阳离子转运系统转运进入细胞质。诱导产生细胞质膜的H+—ATP酶来酸化根周围环境从而提高Fe的供应量。相应基因:Fe(Ⅱ)—转运蛋白(IRT)基因,Fe(Ⅲ)—还原酶(FRO)基因,H+—ATPase酶。机理Ⅱ禾本科植物吸铁:植物铁载体(PS)在细胞质中由甲硫氨酸通过尼克酸胺途径合成,一种铁缺乏诱导的质膜(PM)转运蛋白参与PS释放到根际的过程,质膜中另一种铁缺乏诱导的转运蛋白识别Fe—PS复合物并将其完整地转运至细胞质中。相关基因:S—腺苷甲硫氨酸合成酶基因,尼克烟酰胺合成酶基因,尼克烟酰胺氨基转移酶基因,Fe(Ⅲ)—MAS转运蛋白基因。机理Ⅲ(拟南芥,水稻):相关基因:类似转铁的蛋白,Nramp基因家族。植物对锌的吸收存在两种现象:通过调控植物根系生物学特性来完成锌的吸收;通过诱导表达根部锌吸收转运蛋白来完成,但此过程需消耗能量。锌转运蛋白:ZIP家族主要将锌从细胞各个区室运输进入细胞质溶液中,CDF家族主要将锌从细胞质溶液中排出体外或将细胞质内过量的锌运输进入细胞内的器官储藏起来。P—型ATP酶重金属转运蛋白,该蛋白的存在对降低细胞锌的浓度和锌离子平衡具重要意义。其他锌转运蛋白。锌转运蛋白基因表达差异:锌转运载体的物种和品种特异性;转运蛋白在器官组织和细胞中的特异性;锌转运蛋白的离子选择性、方向性和亲和力;载体蛋白基因的表达受环境的影响。环境中锌离子的浓度影响蛋白的表达,高亲和力的载体蛋白使植物在缺锌或铁的情况下,能够保持一定的锌或铁浓度,以适应生理需要;而低亲和力载体则是对毒害的适应。PH影响蛋白的表达。二、矿质养分离子进入根内部及导管的途径及相关概念。简述膜蛋白种类及各分类作用机理。离子可以通过外质体和共质体途径进入根的内部。由于内皮层上凯氏带的存在,离子只能够通过共质体途径进入中柱,再经过外质体途径扩散进入导管(或管胞)。离子从木质部薄壁细胞释放到导管的过程以受代谢过程控制的主动过程为主,也有被动扩散过程。外质体(apoplastorapoplasm):由细胞壁、细胞间隙和无细胞质的细胞内腔(如木质部导管和纤维)组成的连续系统。在根部,外质体途径被凯氏带所阻断。共质体(symplastorsymplasm):通过胞间连丝将细胞质相互连接组成的整个网络。胞间连丝(plasmodesmata):是沿质膜排列的直径为20-60nm的柱形孔道,其中有连丝微管通过,连丝微管是由两个细胞的光滑内质网衍生而来的。外质体和共质体对流运输的机制:外质体:蒸腾作用提供驱动压力共质体:渗透压产生的压流1)通道(channel)蛋白:膜蛋白,在膜上形成选择性孔道,使分子或离子通过这些孔道扩散过膜。2)离子泵(ionpumporiontransporter)是能逆离子浓度梯度转运离子通过质膜的跨膜蛋白。a.质子泵(protonpump):也称为H+-ATP酶,是一种利用能量(来自ATP水解)逆着电化学梯度转运H+的过程,因此是一个主动运输过程。b.钙泵(calciumpump):也称为Ca2+-ATP酶,它催化质膜内侧的ATP水解释放能量,驱动细胞内的钙离子泵出细胞。3)载体(carrier)蛋白:质膜上的一类内在蛋白,可以选择性地与质膜一侧的分子或离子结合,形成载体—分子(或离子)复合物。再通过载体蛋白构象的变化,透过质膜,把分子或离子释放到质膜的另一侧。可以顺电化学梯度进行(被动运输),但以逆电化学梯度进行为主(主动运输)。单向运输载体(uniportcarrier):顺电化学势的单向运输。同向运输体(symporter):被运输的离子与质子被载体蛋白运向膜的同一侧。如:硝酸、磷酸、硫酸、氨基酸及蔗糖等。反向运输体(antiporter):被运输的离子与质子的运输方向相反,如:K+、Na+等。三、光受体有哪些及PHY的自我磷酸化作用机理光敏受体在植物光反应中,光受体包括:(1)吸收红光/远红光(波长为600~750nm)的光敏色素(phytochrome);(2)吸收蓝光/UV-A(波长为320~500nm)的隐花色素(cryptochrome)和向光素(phototropin);(3)UV-B(波长为282~320nm)受体(UV-Breceptor)蛋白激酶(proteinkinase)是一类催化ATP上的磷酸基团转移到自身或其他蛋白质的丝氨酸或苏氨酸上,调节蛋白质的活性;蛋白磷酸酶(proteinphosphatase)是从蛋白质上移去磷酸基团的酶,可通过拮抗蛋白激酶而调节蛋白质活性。因此,蛋白激酶和蛋白磷酸酶在信号转导中起相反作用。光敏色素是具有自我磷酸化能力的蛋白激酶。然而高等植物的光敏色素并不具有组氨酸激酶活性,而是光调节自磷酸化的苏氨酸/丝氨酸激酶,照射红光后,生色团受光刺激,使C端的蛋白激酶活化,将ATP磷酸基团转移到N端的丝氨酸残基,光敏色素分子因自身磷酸化而活化,而后再使下游的X因子发生磷酸化(图),同时也将红光信号传递给下游的X组分。四、为什么说光敏素的发现是植物科学发展史中的重要事件?抑制PHY在植物个体发育中有哪些作用?举例说明。植物个体发育的整个过程都离不开光敏色素的作用?1、从调节种子萌发到幼苗胚芽鞘、中胚轴生长、幼叶的展开,或下胚轴的生长、弯钩伸直、两片子叶的展开和伸展。2、从叶片表面气孔的形成和活动到幼茎维管束的分化、表皮毛的形成3、从植株地上部发育形态到根的生长和根冠比的调节。4、从叶绿体的发育到光合活性的调控。5、从贮藏淀粉、脂肪的降解到硝酸盐的还原同化,蛋白质、核酸的合成。6、从光周期控制花芽分化到叶片器官衰老的调节1、高等植物中一些由光敏色素控制的反应:1.种子萌发2.弯钩张开3.节间延长4.根原基起始5.叶分化和扩大6.小叶运动7.膜透性8.向光敏感性9.花色素形成10.质体形成11.光周期12.花诱导13.子叶张开14.肉质化15.偏上性16.叶脱落17.块茎形成18.性别表现19.单子叶展叶20.节律现象2、简述PHY的三种反应类型;LFR、RLFR、HIR的作用特点LF(低辐射反应):1μmol/m2的光即可启动这类反应。1000μmol/m2时达到饱和,反应可被一短暂的红闪光有效诱导并可被随后的远红光照射逆转,多数光形态建成是LFRVLFR(极低辐射反应):被10-410-2μmol/m2的红光或远红光诱导,氮红光反应不能被远红光逆转。VLFR仅种子吸涨或幼苗完全生长在黑暗中才能鉴别出来,即使实验室绿色安全光下也可发生反应:VLFR的作用光谱峰在红光和蓝光区,说明其受体是光敏色素,而且可能是有蓝光受体参与。HIR(高辐射反应):需要相对强的连续光照的反应,激活反应的光强需大于10μmol/m2,达到饱和的光强度比LFR高100倍,是波长,辐照强度及光照时间的综合作用。光照越强,光照时间越长,则反应越大。幼苗下胚轴伸长的抑制,弯钩伸直,去黄化,及一些植物花色素苷的合成都是HIR。此类反应的作用光谱在红光、远红光、蓝光/UV-A区,其受体可能有PHY和蓝光受体。3、光敏色素的可能作用机理有哪些?举例说明。三种假说:酶假说:PHY是一种酶,PFR是有活性的形式。有人提出Phy可能是一种受光调节的蛋白激酶,即Phy既是光受体,又是蛋白激酶。相反的观点认为蛋白激酶污染了Phy制品或Phy与蛋白激酶相连造成。膜透性假说:Phy与细胞膜相互作用,转型后影响膜的透性,进而影响例子通透性和酶系活动等各方面。Phy控制的快速反应支持此假说。但缺乏直接的证据说明Phy和膜的作用能诱导某个反应。基因调控假说:越来越多的科学家倾向于这种假说。这个假说能解释Phy的慢反应。需要较长的停滞期,形成各种蛋白质和酶。4、光敏色素信号转导G蛋白Pfr激活G蛋白可能通过下列两种方式:一是可溶性光敏色素转移到膜上直接调节G蛋白活性;二是光敏色素先与膜上另一种蛋白质发生相互作用,然后由后者去调节G蛋白。Ca2+和CaMRoux等提出钙信使系统参与红光信号转导的假说。在动物细胞中,已知钙信使系统通过下列两种方式调节基因的表达:一是间接方式,通过Ca2+/CaM依赖的蛋白激酶将转录因子磷酸化以调节转录活性;二是直接方式,被Ca2+活化的CaM直接与具螺旋-环-螺旋(basic-helix-loop-helix)超二级结构(motif)的转录因子相结合,进而抑制这些转录因子与DNA的结合。cGMPBowler等首次证实cGMP参与了植物红光信号转导。IP3和DG研究表明在光调节的气孔开闭过程中IP3起信号转导作用五、获得次生代谢产物的方法,如何提高次生代谢产量?次生代谢物的合成途径及生理功能?1、获得植物次生代谢物的方法(1)直接从植物材料中提取;(2)用化学合成模拟;(3)真菌或细菌发酵生产;(4)利用植物组织和细胞培养法生产次生物质。2、提高次生代谢产量(1)植物次生代谢基因工程是利用基因工程技术对植物次生代谢途径的遗传特性进行改造,改变植物次生代谢产物。次生代谢基因工程是通过转入某些关键酶的基因或其反义基因,促进或抑制该基因的表达,进而改变植物次生代谢产物。(2)次生代谢物生产的细胞工程直接从野生植株中分离提取植物细胞培养常用的细胞培养技术:①细胞悬浮培养法②两相培养技术③反义技术植物发根培养-毛状根培养技术3、次生代谢物的合成途径(1)、类异戊二烯途径——产生萜类化合物:植物类异戊二烯的生物合成至少存在两种生物合成途径。甲羟戊酸途径(MVA):在细胞质中进行。并一糖酵解产物乙酰辅酶A作为原初供体,合成倍半萜、三萜和甾体类等。丙酮酸/磷酸甘油醛途径(DXP),又称非依赖甲羟戊酸途径:在质体中进行。以丙酮酸和磷酸甘油醛为原料,合成胡萝卜素、单萜和二萜等。(2)、苯丙烷代谢途径——产生酚类化合物:多种途径产生。莽草酸途径:高等植物合成酚类的主要方法。丙二酸途径:真菌和细菌。(3)、氨基酸代谢途径——含N生物碱化合物是由氨基酸转化而来。4、次生代谢物的功能1、生长发育的调控2、提高抗逆性3、抗氧化作用4、信号物质5、低于天敌的侵袭6、增强抗病性7、提高植物的种间竞争能力8、维系植物与其他生物之间的互惠关系六、何为相容性物质(渗透调节),特点,及其作用机理(假说)?简述甜菜碱的合成途径及生理功能。1、相容性物质Compatiblesolutes渗透调节是指细胞渗透势变化所表现出的调节作用。通过细胞增加或减少溶质调节细胞渗透势以期与细胞外渗透势平衡。植物细胞吸收的无机离子,K+,Na+,Ca2+,Cl-,等细胞内合成的有机溶质:脯氨酸,甜菜碱,糖类(山梨醇,甘露醇),有机酸等。相容性物质的特点:在生理pH范围内不带净电荷;能为细胞膜所保持;引起酶变化的作用最小;产生迅速。分子量小,易溶于水。2、作用机制——两种主要假说表面结合假说:相容性物质与蛋白质表面特别是疏水表面结合,提高蛋白质表面水化程度。蛋白质表面的疏水区域是非常脆弱或敏感的部分,因为结合在这里的水分子结合能力非常弱,细胞在水分胁迫环境中将首先脱去这部分水。相容性物质与蛋白质疏水区域结合后,将其转化为亲水表面。溶质排除假说:在蛋白质溶液中,相容性物质将首先溶解在大量的自由水中从而提高蛋白质分子表面的水化程度,而不是溶在蛋白质或其他生物大分子表面的结合水中,不参与生物大分子的直接作用。3、甜菜碱:是一类季铵化合物,化学名称为N-甲基代氨基酸。植物中的甜菜碱主要有12种,研究最多的是甘氨酸甜菜碱生物合成:甜菜碱是以胆碱为底物经两部酶催化生成,即:胆碱――(1)→甜菜碱醛――(2)―→甜菜碱在高等植物中,甜菜碱由胆碱单氧化酶(CMO)和甜菜碱醛脱氢酶(BADH)催化合成。在哺乳动物及微生物,如大肠杆菌细胞中,甜菜碱由胆碱脱氢酶(CDH)和甜菜碱醛脱氢酶(BADH)催化合成
 三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
三七文档所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 扫描二维码
扫描二维码








 neozoo
neozoo
本文标题:高级植物生理复习.总结
链接地址:https://www.777doc.com/doc-4740140 .html